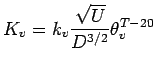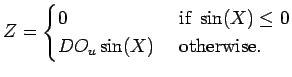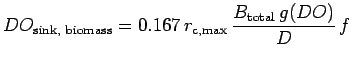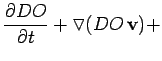 div div |
(A.8) |
The Neuse is characterized as an intermittently-mixed estuary with wind being the primary mechanism controlling vertical mixing (Borsuk et al. 2001b; Luettich Jr. et al. 1999) temporarily dissipating bottom water hypoxia. Hypoxia is typically defined as dissolved oxygen concentrations (DO) ranging from 0.2 to 2 mg/L with anoxic conditions occurring below 0.2 mg/L. Over an entire year, DO in the Neuse ranges from 0.1 to 10.6 mg/L with hypoxic conditions developing intermittently during summer, lasting from a few days to weeks and varying intra and inter seasonally (Selberg et al. 2001). Empirically, the duration of continuous hypoxia at a given location is about 9 days in the deep parts of the estuary which experience multiple hypoxic episodes throughout the summer. The primary mechanisms causing hypoxia at any location in the estuary is oxygen usage by the biomass and sediments (which is temperature dependent) while the primary mechanisms dissipating hypoxia are oxygen diffusion from the surface waters (which is also a temperature dependent process) and random wind events that mix the oxygen rich surface waters with the oxygen depleted bottom waters.
The model for DO (mg/L) results from greatly simplifying a more complex model (Mauersberger 1983) obtained using the principle of mass conservation:
Because the Neuse has a very long residence time (Kim 1990; Luettich Jr. et al. 1999), the advection term,
![]() , is
considered insignificant. Under calm conditions that favor the
formation of hypoxia,
, is
considered insignificant. Under calm conditions that favor the
formation of hypoxia,
![]() is dominated by molecular
diffusion which is small (Hyer et al. 1971) and thus will be
considered insignificant. The overall sink term is split into two
components: oxygen demand from sediments and demand from clam and
background biomass. Similarly, the source term is split into a
component dealing with re-aeration from the surface and a second
component,
is dominated by molecular
diffusion which is small (Hyer et al. 1971) and thus will be
considered insignificant. The overall sink term is split into two
components: oxygen demand from sediments and demand from clam and
background biomass. Similarly, the source term is split into a
component dealing with re-aeration from the surface and a second
component, ![]() , dealing with random mixing events (i.e., wind) and is
used to control the shape, duration and extent of hypoxia in the model
estuary. This yields:
, dealing with random mixing events (i.e., wind) and is
used to control the shape, duration and extent of hypoxia in the model
estuary. This yields:
Random mixing events, ![]() (mg.L
(mg.L![]() .hr
.hr![]() ) in
Eqn A.9, are modeled by transforming a Gaussian random
field. The Gaussian field used to generate bottom salinity,
) in
Eqn A.9, are modeled by transforming a Gaussian random
field. The Gaussian field used to generate bottom salinity, ![]() , is
used (Appendix A.3.3), after it is transformed by:
, is
used (Appendix A.3.3), after it is transformed by:
The rate at which oxygen is used depends on the total clam and
background biomass,
![]() total (g/m
total (g/m![]() ) present on a triangle
and the oxygen usage by the sediments. Oxygen utilization by crabs is
ignored since crabs are mobile and crab biomass density per m
) present on a triangle
and the oxygen usage by the sediments. Oxygen utilization by crabs is
ignored since crabs are mobile and crab biomass density per m![]() is
much less than the combined clam and background biomass density. It
is assumed that clam and background biomass can be combined into one
term. The rate of oxygen usage (mg.L
is
much less than the combined clam and background biomass density. It
is assumed that clam and background biomass can be combined into one
term. The rate of oxygen usage (mg.L![]() .hr
.hr![]() ) by the biomass
on a triangle is similar to Hummel (1985a):
) by the biomass
on a triangle is similar to Hummel (1985a):
The oxygen used by the sediments (mg.L![]() .hr
.hr![]() ) on the
triangle is given by:
) on the
triangle is given by:
For a triangle at a depth of 5 m, a DO of 4 (mg/L) and temperatures of
20, 25 and 30
![]() sediment oxygen demand is
sediment oxygen demand is
![]() , 0.053,
0.078 (mg.L
, 0.053,
0.078 (mg.L![]() .hr
.hr![]() ), respectively and if the density of
biomass on the triangle is 800 g/m
), respectively and if the density of
biomass on the triangle is 800 g/m![]() , its oxygen demand is
, its oxygen demand is ![]() 0.009, 0.007 and 0.002 (mg.L
0.009, 0.007 and 0.002 (mg.L![]() .hr
.hr![]() ) respectively, since the
biomass uses less oxygen at higher temperatures. The rate of
re-aeration of the bottom waters depends on depth
(Eqns A.10 and A.12) with deeper waters taking
longer to re-aerate than shallower. Following mixing, deeper waters
take longer to become hypoxic because more oxygen is stored above
them. Finally, in Eqn (A.9), we also add the
constraints that DO
) respectively, since the
biomass uses less oxygen at higher temperatures. The rate of
re-aeration of the bottom waters depends on depth
(Eqns A.10 and A.12) with deeper waters taking
longer to re-aerate than shallower. Following mixing, deeper waters
take longer to become hypoxic because more oxygen is stored above
them. Finally, in Eqn (A.9), we also add the
constraints that DO ![]() and DO
and DO ![]() .
.
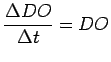 source
source