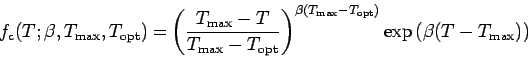Next: Clam Mortality
Previous: Models for Clams and
Clam Growth Dynamics
Adult clams are generally less than 2.5 cm
wide (McErlean 1964), although a few may reach 3.0 cm (Bachelet 1980). In northern climates, clams can live as long
as 35 years (Bachelet 1980), while 6-10 years is common in more
southern climates (Gilbert 1973). Clam growth is most rapid
between April and June (1.9-3.7 mm/month) (Holland et al. 1987),
slows in July and August and continues to decrease throughout the
fall/winter. The variability in clam growth rates depends on
temperature and possibly salinity resulting in southern clams growing
faster than those further north. For Rand Harbor, Massachusetts,
clams grew 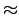 1.0 and
1.0 and 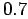 cm in the first two years,
respectively, before growth slowed
considerably (Gilbert 1973). A similar growth pattern has been
observed in other locations (Commito 1982; Bachelet 1980). At
temperate latitudes, the mass of bivalve molluscs generally declines
during winter, declining more during warmer than colder
winters (Honkoop and Beukema 1997). Females reproduce when they are
greater than 1 cm, or approximately 2.9 yrs for clams in northern
climates like Maine (Commito 1982) and at a younger age in
warmer climates.
cm in the first two years,
respectively, before growth slowed
considerably (Gilbert 1973). A similar growth pattern has been
observed in other locations (Commito 1982; Bachelet 1980). At
temperate latitudes, the mass of bivalve molluscs generally declines
during winter, declining more during warmer than colder
winters (Honkoop and Beukema 1997). Females reproduce when they are
greater than 1 cm, or approximately 2.9 yrs for clams in northern
climates like Maine (Commito 1982) and at a younger age in
warmer climates.
Our clam growth model closely follows the bivalve model proposed by Solidoro et al. (2000). We assume that clams on each triangle
grow according to the same model with differences in growth rate
occurring between triangles due to spatial differences in temperature
and dissolved oxygen. Possible salinity effects are
ignored (Holland et al. 1987). Because anthropogenic loading should
increase algae, the primary food of clams (Hummel 1985b), we
assume that clam growth is not limited by food
availability (Holland et al. 1980). Clam wet weight,
 c,w (g), is related to shell length,
c,w (g), is related to shell length,
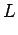 c (cm), by
the allometric model
c (cm), by
the allometric model
where
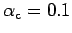 g.cm
g.cm
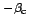 and
and
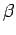 c
c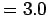 (Bachelet 1980; Gilbert 1973, Table 2).
The maximum size of clams in the Neuse is 2.5
cm (Bachelet 1980, Table 7).
(Bachelet 1980; Gilbert 1973, Table 2).
The maximum size of clams in the Neuse is 2.5
cm (Bachelet 1980, Table 7).
Clam growth rate is proportional to the difference in the grams of energy
available for feeding and the energy used for
respiration (Solidoro et al. 2000, Eqn 1):
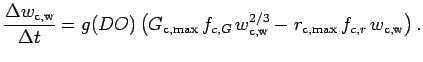 |
(A.17) |
The temperature and DO values used are constant over 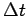 and
the values used are those from the start of the time interval.
and
the values used are those from the start of the time interval.
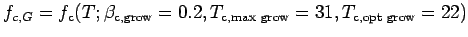 and
and
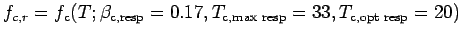 are given by Eqn (A.18) and
are given by Eqn (A.18) and 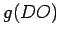 by
Eqn (A.19).
by
Eqn (A.19).
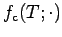 is maximum at the
optimal temperature and alters clam intake and respiration with bottom
water temperature. The rate of clam growth at a given temperature and
mass is shown in Fig. A4.
is maximum at the
optimal temperature and alters clam intake and respiration with bottom
water temperature. The rate of clam growth at a given temperature and
mass is shown in Fig. A4.
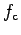 is given by:
is given by:
which is taken from Solidoro et al. (2000) and is similar in shape
to that found by McMahon and Wilson (1981, Fig. 2).
Bivalves either conform to or regulate their rate of oxygen
consumption independent of oxygen
concentrations (Wang and Widdows 1993). In Macoma, oxygen
consumption decreased with decreasing DO
concentration (McMahon and Wilson 1981, Fig 5) and based on these results, we
assume that this decrease in oxygen consumption represents an overall
decrease in growth rate in Eqn (A.17) according to:
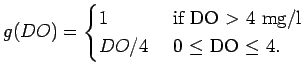 |
(A.19) |
Thus, decreases linearly with DO for DO
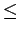 4 mg/L.
4 mg/L.
The wet weight of the clams in each of the 8 year classes for spring
and fall recruitment on each triangle in the estuary are updated
according to Eqn (A.17) every 24 hours. The maximum
rate of respiration,
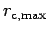 =0.00034 (1/hour), is based on McMahon and Wilson (1981) and Hummel (1985a, Table 1). In
Eqn (A.17), as a clam's mass increases its respiratory
demands increase faster than its rate of food intake making it
impossible for a clam to grow beyond a given mass. This fact was used
to establish
=0.00034 (1/hour), is based on McMahon and Wilson (1981) and Hummel (1985a, Table 1). In
Eqn (A.17), as a clam's mass increases its respiratory
demands increase faster than its rate of food intake making it
impossible for a clam to grow beyond a given mass. This fact was used
to establish
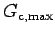 at 0.00034 (g
at 0.00034 (g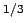 /hr).
/hr).
Next: Clam Mortality
Previous: Models for Clams and