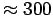
Clams are iteroparous broadcast spawners, with males and females
releasing gametes synchronously, typically between May to June
although fall recruitment can also occur depending on how far south
the clams are located (Harvey and Vincent 1989; Gilbert 1978). Release
of sperm and eggs is likely triggered by temperature cues (7 to
14
![]() ), spring tides and/or quantity of food
available (Harvey and Vincent 1989; Gilbert 1978). After a few weeks of
pelagic life, the larvae (
), spring tides and/or quantity of food
available (Harvey and Vincent 1989; Gilbert 1978). After a few weeks of
pelagic life, the larvae (
![]() m) settle from the water
column to the sediment (Bouma et al. 2001). Juvenile clams are
capable of migration (Beukema and de Vlas 1989), but this is unlikely to
be an important factor in spatially structuring clam distribution.
m) settle from the water
column to the sediment (Bouma et al. 2001). Juvenile clams are
capable of migration (Beukema and de Vlas 1989), but this is unlikely to
be an important factor in spatially structuring clam distribution.
Using a mesh size of 0.5 mm, Hines and Comtois (1985) reported 34,000
clams per m (119 g/m dry weight, 780 g/m wet weight) in
mud and 22,000 clams per m (177 g/m dry weight, 1150 g/m wet weight) in sand at the mouth of the Rhode River, Maryland -
similar to the biomass and larval densities reported for sites in
Europe (Bouma et al. 2001; Rosenberg et al. 1992). In the Dutch Wadden
Sea, recruit to egg ratio for clams varied between 0.0001 to
0.001 (Honkoop et al. 1998). Although 37% of year-to-year
variation in recruit densities could be explained by inter-annual
variation in winter temperature, only 7% was explained by variation
in egg density. Thus, the number of adults and total spawned eggs are
poor predictors of subsequent recruit
abundance (Honkoop et al. 1998).
(119 g/m dry weight, 780 g/m wet weight) in
mud and 22,000 clams per m (177 g/m dry weight, 1150 g/m wet weight) in sand at the mouth of the Rhode River, Maryland -
similar to the biomass and larval densities reported for sites in
Europe (Bouma et al. 2001; Rosenberg et al. 1992). In the Dutch Wadden
Sea, recruit to egg ratio for clams varied between 0.0001 to
0.001 (Honkoop et al. 1998). Although 37% of year-to-year
variation in recruit densities could be explained by inter-annual
variation in winter temperature, only 7% was explained by variation
in egg density. Thus, the number of adults and total spawned eggs are
poor predictors of subsequent recruit
abundance (Honkoop et al. 1998).
The model uses two discrete spawning events which occur on March 31 and August 31. At these times, female clams (assumed to be half of the population) with a shell length greater than 1 cm reproduce (Commito 1982; Honkoop et al. 1998). The number of eggs produced per female is:
), is the total number
of eggs divided by the area of the estuary and multiplied by a random
estuary recruitment factor which is generated uniformly over the range
0.0001 to 0.001 (Honkoop et al. 1998). How the potential
recruitment egg density is altered on each triangle by depth, current
clam biomass and density is described below.
It is necessary to control the density of clam egg recruitment on each
triangle because our model does not include all of the different
predators and causes of mortality present in the actual estuary. In
summary, fewer spat are assumed to settle on shallower (![]() 1.5 m
deep) than deeper areas and recruitment is limited both by a
triangle's current clam biomass density
(Eqn A.23) and clam density
(Eqn A.24).
1.5 m
deep) than deeper areas and recruitment is limited both by a
triangle's current clam biomass density
(Eqn A.23) and clam density
(Eqn A.24).
Let
![]() be the depth factor limiting clam
recruitment on shallow triangles and let
be the depth factor limiting clam
recruitment on shallow triangles and let ![]() (g/m) be the
grams of clam biomass on the triangle and
(g/m) be the
grams of clam biomass on the triangle and
![]() the factor
limiting clam recruitment on triangles with high clam biomass:
the factor
limiting clam recruitment on triangles with high clam biomass:
(Hines and Comtois 1985).
Finally, let the density of clams already present on each triangle be
given by
![]() (#/m) and let
(#/m) and let
![]() (#/m) (Holland et al. 1980) be an upper bound on clam density.
Accounting for depth, current clam biomass and clam density, the
number of clams recruited to a triangle as a function of
(#/m) (Holland et al. 1980) be an upper bound on clam density.
Accounting for depth, current clam biomass and clam density, the
number of clams recruited to a triangle as a function of ![]() ,
(or potential recruitment egg density), is:
,
(or potential recruitment egg density), is:
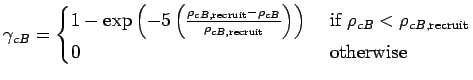
![$\displaystyle ) = \begin{cases}\left((\rho_{c u} - \rho_\text{c,tri})+\rho_{cu}...
...ght)^2\right]} & \text{ if } \rho_{\text{c,tri}} \geq \rho_{c u} \\ \end{cases}$](img138.png)