Ecological Archives E096-185-A2
Nitin Sekar, Chia-Lo Lee, and Raman Sukumar. 2015. In the elephant's seed shadow: the prospects of domestic bovids as replacement dispersers of three tropical Asian trees. Ecology 96:20932105. http://dx.doi.org/10.1890/14-1543.1
Appendix B. Seasonality and sex-based differences in movement data.
D. indica fruits predominantly from December through April, and C. arborea and A. chaplasha fruit from late June through early August. Thus, movement data for our animal study species should ideally be partitioned based on season to capture differences in how the study species move during different parts of the year. Unfortunately, since we have limited movement data, we are more likely to lose important variation in movement patterns between different individuals/herds than to capture meaningful interseasonal differences by subsetting the data.
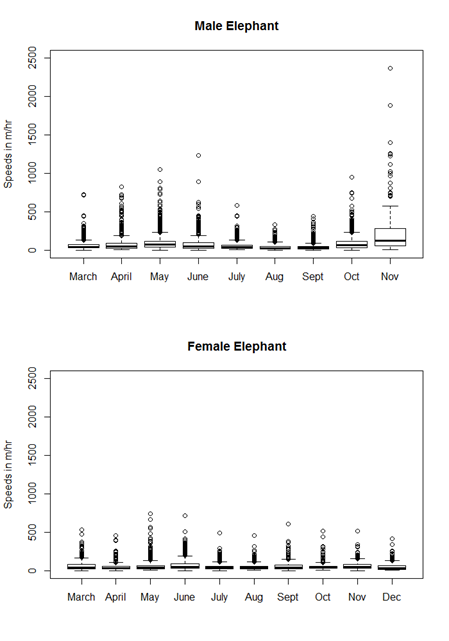
As noted in the main text, we have detailed movement data for two elephants, one male and one female, from March to November/December. In Fig. B1 below, we plot boxplots of the average speeds calculated for each individual every 8 hours (same starting point, with ending points every 8 hours) during each month of the study. While we did not use speed directly in our seed displacement simulation, the distribution of speeds faithfully shows how elephant movements (and thus our calculations of seed displacements) varied amongst months.
Several points are worthy of note. First, due to the timing of the data collected, we miss much of the D. indica fruiting season (there is no data for mid-December through February) for both elephants. Secondly, it appears that the male elephant sometimes moves substantially faster than the female elephant, likely because the latter is constrained by her family group. This is especially true in November, when the male elephant is calculated to move between 1 and 2.5 km an hour for hours or days at a time. These speeds are responsible for the greatest seed dispersal distances found in our study. It is possible that the elephant was in musth at this time; it is well known that adult male elephants increase their movement rate and home range size substantially when they come into musth (Fernando et al. 2008), and in the Buxa population males do experience musth during the fruiting seasons as well (personal observation). Thirdly, it appears that both the male and elephants move at somewhat greater speeds in May and June and somewhat less in August.
Fig. B1. Speeds in m/hr calculated for the two GPS-collared elephants from March through November/December of 2005. The male appears to be able to move faster than the female, especially in November (when he was apparently in musth). There is possibly some seasonality in elephant movements, as both individuals often move faster in May and June.
These observations present two possible biases. First, if females and family herds move shorter distances than males and most fruits are eaten by members of family herds, our data and analyses may over-represent seed dispersal by fast- and far-moving male elephants. Second, there is the possibility of seasonal fluctuations in movement with greater movement in May and June than in much of the rest of the year. Since none of our study species fruit in May, this could mean that we are somewhat overestimating the frequency longer-distance dispersal events.
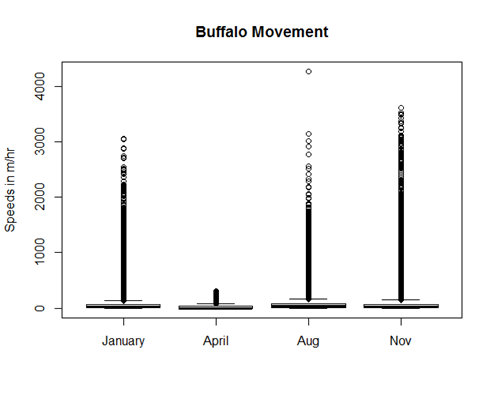
For bovids, movement data was even more limited. In the case of buffaloes, for instance, we followed three different groups over four different periods throughout the year; differences in movement are likely to have more to do with the group at hand (perhaps the number of buffaloes, whether they have a cowherd, or the part of Buxa in which they live) than the season of tracking. In Fig. B2, the April group moves the least, but this likely has more to do with the fact that that herd was feral than the time of year.
Fig. B2. Speeds in m/hr calculated for the four buffalo groups over the 7–10 day periods during which they were followed. The much lower speeds of the April group likely reflects the fact that the group was feral as opposed to the season in which measurements were taken.
Overall, our hope is that our movement data has captured enough of the variation in movement patterns amongst individuals or groups of a given species to allow for reasonable comparisons in dispersal ability.
Literature cited
Fernando, P., E. D. Wikramanayke, H. K. Janaka, L. K. A. Jayasinghe, M. Gunawardena, S. W. Kotagama, … and J. Pastorini. 2008. Ranging behavior of the Asian elephant in Sri Lanka, 73(1):2–13.