Volume 88, Number 1, January 2007
Volume
88, Number 1, January 2007
 Cover
Photo: The consequences of climate change for ecosystem structure and function remain largely unknown. A 20-year study of a high-elevation riparian system in Arizona shows substantial changes in both structure and function related to climate change. Seven species of birds were found to initiate earlier breeding associated with an increase in spring temperature across years. Climate had much larger consequences for these species by affecting trophic levels below (plants) and above (predators) them. The Orange-crowned Warbler (Vermivora celata ) depends on maple for nesting and feeding. It has shown a major decline in abundance over the 20 years, associated with the decline in maple, but also associated with decreasing summer precipitation and increasing nest predation rates.
Cover
Photo: The consequences of climate change for ecosystem structure and function remain largely unknown. A 20-year study of a high-elevation riparian system in Arizona shows substantial changes in both structure and function related to climate change. Seven species of birds were found to initiate earlier breeding associated with an increase in spring temperature across years. Climate had much larger consequences for these species by affecting trophic levels below (plants) and above (predators) them. The Orange-crowned Warbler (Vermivora celata ) depends on maple for nesting and feeding. It has shown a major decline in abundance over the 20 years, associated with the decline in maple, but also associated with decreasing summer precipitation and increasing nest predation rates.
See the Photo Gallery for other photographs associated with this article by Thomas E. Martin.
Visit the Photo Gallery for more photographs submitted by our scientific journal authors.
Table of Contents
(click on a title to view that section)
FROM THE EDITOR-IN-CHIEF
Request for an Editor: New Web Sites
ANNOUNCEMENTS
Society Notices
Results of Elections for ESA Offices
Call for Nominations: ESA Honorary Member Award
Student Awards for Excellence in Ecology
Call for Student Award Judges
Other Notices
Vegetation Classification Draft Standard
Resolution of Respect: Frank B. Golley
LETTERS TO THE BULLETIN Society Actions
Minutes of the 5–6 August Governing Board Meeting
Minutes of the 11 August Governing Board Meeting
PHOTO GALLERY: Images from upcoming articles in our scientific journals
Littorine Snails Compete by Altering a Shared Resource. J. Long, R. S. Hamilton, and J. L. Mitchell
Trophic Changes in a High-elevation Riparian System. T. E. Martin
Thermal Stress in the Rocky Intertidal Zone. B. Helmuth, B. R. Broitman, C. A. Blanchette, S. Gilman, P. Halpin, C. D. G. Harley, M. J. O’Donnell, G. E. Hoffman, B. Menge, and D. Strickland
Mangrove Forest Structure. W. P. Sousa, P. G. Kennedy, B. J. Mitchell, and B. M Ordóñez L.
CONTRIBUTIONS
Commentary
Understanding Food Chains and Food Webs. F. N. Egerton
Advocacy and Integrity. S. Sorooshian
A History of the Ecological Sciences, Part 23. Linnaeus and the Economy of Nature. F. N. Egerton
Click here for a full complement of this series by Frank N. Egerton
DEPARTMENTS
Public Affairs Perspective
Rapid Response Team Fall Newsletter
ESA Statement on Global Climate Change
Ecology 101
Preparing for the Field Season. C. Wong
The Paper Trail
Essay on William S. Cooper’s “The Fundamentals of Vegetational Change.” S. T. A. Pickett
Reports of Symposia at the ESA Annual Meeting
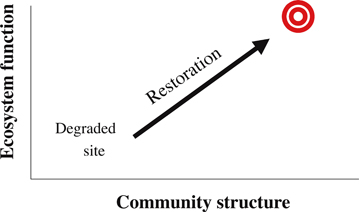
Upstart View of Restoration Icons. J. B. Zedler, D. A. Falk, and D. J. Larkin
Urban Food Webs: Errata: Revised text and figure. P. Warren et al.
Society Section and Chapter News
Canada Chapter Newsletter
Southeastern Chapter Newsletter
Meetings
Meeting Calendar
Urban Wildlife Management National Conference
The BULLETIN OF THE ECOLOGICAL
SOCIETY OF AMERICA (ISSN 0012-9623)
is published quarterly by the
Ecological Society of America, 1707 H Street, N.W., Suite 400, Washington, DC
20006.
It is available online only, free of charge, at ‹http://www.esapubs.org/bulletin/current/current.htm›.
Issues published prior to January 2004 are available through
‹http://www.esapubs.org/esapubs/journals/bulletin_main.htm›
Bulletin
of the Ecological Society of America, 1707 H Street, NW, Washington DC 20006
Phone (403) 220-7635, Fax (403) 289-9311,
E-mail: [email protected]
Associate
Editor Section
Editor, Ecology 101 |
Section
Editors, |
The
Ecological Society of America
GOVERNING BOARD FOR 2006–2007
President: Alan Covich, Institute of Ecology, University of Georgia, Athens, GA 30602
President-Elect: Norm Christensen, Nicholas School of the Environment and Earth Sciences, Duke University, Durham, NC 27708
Past-President: Nancy B. Grimm, School of Life Sciences, Arizona State University, Tempe, AZ 85287-4501
Vice President for Science: Gus R. Shaver, The Ecosystems Center, Marine Biological Laboratory, Woods Hole, MA 02543
Vice President for Finance: William J. Parton, Natural Resource Ecology Laboratory, Colorado State University, Ft. Collins, CO 80523-1499
Vice President for Public Affairs: Richard V. Pouyat, 3315 Hudson St., Baltimore, MD 21224
Vice President for Education and Human Resources: Margaret D. Lowman, Biology and Environmental Studies, New College of Florida, Sarasota, FL 34243-2109
Secretary: David W. Inouye, Department of Biology, University of Maryland, College Park, MD 20742-4415
Member-at-Large: Dennis Ojima, Natural Resource Ecology Laboratory, Colorado State University, Ft. Collins, CO 80523-1499
Member-at-Large: Jayne Belnap, USGS Cayonlands Field Station, Southwest Biological Science Center, Moab, UT 84532
Member-at-Large: Juan J. Armesto, Departmento de Biologia, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
AIMS
The Ecological Society of
America was founded in 1915 for the purpose of unifying the sciences of
ecology, stimulating research in all aspects of the discipline, encouraging
communication among ecologists, and promoting the responsible application
of ecological data and principles to the solution of environmental problems.
Ecology is the scientific discipline that is concerned with the relationships
between organisms and their past, present, and future environments. These
relationships include physiological responses of individuals, structure
and dynamics of populations, interactions among species, organization
of biological communities, and processing of energy and matter in ecosystems.
| Regular member: | Income level | Dues |
| <$40,000 | $50.00 | |
| $40,000—60,000 | $75.00 | |
| >$60,000 | $95.00 | |
|
Student member:
|
$25.00 | |
| Emeritus member: | Free | |
|
Life
member:
|
Contact Member and Subscriber Services (see below) |
Ecological
Applications $50.00 $40.00
Frontiers in Ecology Free to members
Ecological Archives Free
|
I would like to add a new Department to the Bulletin of the Ecological Society of America featuring web sites that would be of interest to the widest number of ecologists. The Editor of this Department will submit for each Bulletin several web sites and a brief (one paragraph or less) commentary on each site. If you would like to be considered for this position, please send me a CV and an example of how you think the Department could be set up. E. A. Johnson E-mail: [email protected] |
|
ESA 2007 Election Results
|
|
President Vice President for Science Secretary (August 2007–August 2010)
|
Member-at-Large Board of Professional Certification |
Call for Nominations: ESA Honorary Member Award
|
|
Murray F. Buell Award and E. Lucy Braun Award Murray F. Buell had a long and distinguished record of service and accomplishment in the Ecological Society of America. Among other things, he ascribed great importance to the participation of students in meetings and to excellence in the presentation of papers. To honor his selfless dedication to the younger generation of ecologists, the Murray F. Buell Award for Excellence in Ecology is given to a student for the outstanding oral paper presented at the ESA Annual Meeting. E. Lucy Braun, an eminent plant ecologist and one of the charter members of the Society, studied and mapped the deciduous forest regions of eastern North America and described them in her classic book, The Deciduous Forests of Eastern North America. To honor her, the E. Lucy Braun Award for Excellence in Ecology is given to a student for the outstanding poster presentation at the ESA Annual Meeting. A candidate for these awards must be an undergraduate, a graduate student, or a recent doctorate not more than 9 months past graduation at the time of the meeting. The paper or poster must be presented as part of the program sponsored by the Ecological Society of America, but the student need not be an ESA member. To be eligible for these awards the student must be the sole or senior author of the oral paper (Note: symposium talks are ineligible) or poster. Papers and posters will be judged on the significance of ideas, creativity, quality of methodology, validity of conclusions drawn from results, and clarity of presentation. While all students are encouraged to participate, winning papers and posters typically describe fully completed projects. The students selected for these awards will be announced in the ESA Bulletin following the Annual Meeting. A certificate and a check for $500 will be presented to each recipient at the next ESA Annual Meeting. If you wish to be considered for either of these awards at the 2007 Annual Meeting, you must send the following to the Chair of the Student Awards Subcommittee: (1) the application form below, (2) a copy of your abstract, and (3) a 250-word or less description of why/how the research presented will advance the field of ecology. Because of the large number of applications for the Buell and Braun awards in recent years, applicants may be pre-screened prior to the meeting, based on the quality of the abstract and this description of the significance of their research. The application form, abstract, and research justification must be sent by mail, fax, or e-mail (e-mail is preferred; send e-mail to [email protected]) to the Chair of the Student Awards Subcommittee: Dr. Anita L. Davelos Baines, Dept. of Biology, The University of Texas-Pan American, 1201 W. University Drive, Edinburg, TX 78541-2999 USA. If you have questions, write, call (956) 380-8732, fax (956) 381-3657, or e-mail: [email protected]. You will be provided with suggestions for enhancing a paper or poster. The deadline for submission of form and abstract is 1 March 2006; applications sent after 1 March 2006 will not be considered. This submission is in addition to the regular abstract submission. Buell/Braun participants who fail to notify the B/B Chair by 1 May of withdrawal from the meeting will be ineligible, barring exceptional circumstances, for consideration in the future. Electronic versions of the Application Form are available on the ESA web site, or you can send an e-mail to [email protected] and request that an electronic version be sent to you as an attachment.
Current Mailing Address _____________________________________________________________________________ Current Telephone _________________________________________________________________________________ E-mail __________________________________________________________________________________________ College/University Affiliation ___________________________________________________________________________ Title of Presentation _________________________________________________________________________________ Presentation: Paper (Buell Award) ______ Poster (Braun Award) _______ At the time of presentation I will be (check one): I will be the sole ____ /senior ____ author (check one) of the paper/poster. Signed (electronic signatures are OK) Please attach a copy of your abstract and 250-word or less description of why/how the research presented will advance the field of ecology. |
REQUEST FOR STUDENT AWARD JUDGES Murray F. Buell Award Judges are needed to evaluate candidates for the Murray F. Buell Award for the outstanding oral presentation by a student and the E. Lucy Braun Award for the outstanding poster presentation by a student at the Annual ESA Meeting at Memphis, Tennessee in 2006. We need to provide each candidate with at least four judges competent in the specific subject of the presentation. Each judge is asked to evaluate 3–5 papers and/or posters. Current graduate students are not eligible to judge. This is a great way to become involved in an important ESA activity. We desperately need your help! Please complete and send this form by mail, fax, or e-mail to the Chair of the Student Awards Subcommittee:Dr. Anita L. Davelos Baines, Dept. of Biology, The University of Texas–Pan American, 1201 W. University Drive, Edinburg, TX 78541-2999 USA. (956) 380-8732, fax (956) 381-3657, or e-mail: [email protected]. If you have judged in the past several years, this information is on file. If you do not have to update your information, simply send me an e-mail message, “Yes, I can judge this year.” Name ______________________________________________________________________________________________ Areas of expertise (check all that apply): Provide a few key words or phrases that describe your interests and expertise: _________________________
|
Other NoticesVegetation Classification Panel and the Federal Geographic Data Committee Vegetation Subcommittee The Vegetation Classification Panel of the Ecological Society of America has been working with the Federal Geographic Data Committee Vegetation Subcommittee for several years to revise the established standards for vegetation classification, and add a mechanism for inclusion of the finest levels of the classification hierarchy (Associations and Alliances). A new draft federal standard (to replace and augment the 1997 standard) is now being circulated to the federal agencies for comment. Your comments would also be appreciated. I will compile any comments I receive and forward them to FGDC. I attach an executive summary by Panel member Don Faber-Langendoen. The full draft standard may be seen at ‹http://www.bio.unc.edu/faculty/peet/lab/temp/FGDCDraftStd_Aug2006.pdf› The key things to note are: 1) The upper levels of the hierarchy have been greatly modified. 2) Cultural vegetation is pulled out at the highest level. 3) The whole classification is now shifting to a dynamic content standard (a first for FGDC), where types can be added, deleted, or changed through a peer-review process without going through the entire FGDC agency approval process. ESA is likely to participate in that peer review, though the standard does not include the implementation plan. NATIONAL VEGETATION CLASSIFICATION STANDARD, VERSION 2— WORKING DRAFT Vegetation Subcommittee Federal Geographic Data Committee SUMMMARY – NatureServe version 31 August 2006 INTRODUCTION The United States Federal Geographic Data Committee (hereafter called the FGDC) is tasked to develop geospatial data standards that will enable sharing of spatial data among producers and users. In 1997, the FGDC Vegetation Subcommittee developed a National Vegetation Classification (NVC) Standard (FGDC-STD-005-1997). This document is a proposed revision of the 1997 standard and, if approved, will replace that document. The standard requires that vegetation types be defined and characterized using appropriate data. New vegetation types will be defined and previously defined types will be refined as data continue to be collected, analyzed, and correlated over time. This process is referred to as successive refinement (or successive approximation). Managing the vegetation classification (content standard) dynamically as the classification process is implemented will allow development of the national vegetation classification system (i.e. data classification standard) to proceed efficiently. The overall purpose of this National Vegetation Classification Standard (hereafter referred to as the “Standard”) is to support the development and use of a consistent national vegetation classification (hereafter referred to as the “NVC”) in order to produce uniform statistics about vegetation resources across the nation, based on vegetation data gathered at local, regional, or national levels. This will facilitate cooperation on vegetation management issues that transcend jurisdictional boundaries. It is therefore important that, as agencies map or inventory vegetation, they collect enough data to translate it for national reporting, aggregation, and comparisons. The ability to crosswalk other vegetation classifications and map legends to the NVC will facilitate the compilation of regional and national summaries. This Standard establishes national procedures for classifying existing vegetation for the United States and its Trust Territories that shall be used by Federal agencies to share vegetation information and report national statistics. Existing vegetation is the plant cover, or floristic composition and vegetation structure, documented to occur at a specific location and time. Each Federal agency is free to develop vegetation classification systems that meet their own information and business needs. The ecological characteristics of such local vegetation types can help guide the design of map legends (sets of map units) to address varying land management issues at multiple spatial scales. The NVC is expected to provide the common link to compare and relate these various map legends to each other and facilitate information sharing between federal agencies and other organizations. SUMMARY OF MAJOR CHANGES COMPARED TO THE 1997 STANDARD The structure of the revised NVC hierarchy is a substantial revision of the 1997 hierarchy. The revised hierarchy addresses the following issues, among others:
Examples of how the NVC can be linked to various land cover classification schemes are also provided. OUTLINE OF CURRENT HIERARCHY
Table 1. Major categories used to organize the hierarchy, and the set of Level 1 units proposed to replace the current Level 1 units.
Table 2. Revised hierarchy for natural vegetation with an example.
Table 3. Revised hierarchy for cultural vegetation with examples (from National Resources Inventory).
SUMMARY OF STANDARDS FOR DESCRIPTION AND CLASSIFICATION OF NATURAL VEGETATION Describes how to collect field plot data for vegetation classification purposes. Also provides guidelines for using literature and other data sources in addition to plot data. Classification and description Decribes how to prepare vegetation plot data or literature-based data, how to analyze and interpret the data, build a description of a type, including nomenclature. Peer review of proposed vegetation types Describes how to submit a vegetation type for peer review to an FGDC sponsored Peer Review team. Data management and dissemination Describes the databases, meta data standards, and web sites needed to maintain vegetation classification plot data and NVC type descriptions. SUMMARY OF STANDARDS FOR DESCRIPTION AND CLASSIFICATION OF CULTURAL VEGETATION At this time the critical components for describing and classifying cultural vegetation are not well-developed. It is suggested that, for Agricultural Vegetation, the National Resource Inventory classification be a starting point for the NVC standard, and that where desirable, the standards developed for natural vegetation can be applied. NEXT STEPS Based on comments received on this document (due 6 October), the FGDC vegetation subcommittee will revise the standards document and submit it for official approval to the FGDC Standards Working Group (SWG). The SWG is charged with conducting a public review of the proposed revised standard, and, provided comments and corrections are incorporated to the satisfaction of the SWG, would move to adopt the standard. At the same time, the FGDC vegetation subcommittee with partners to solicit funds to maintain the standard, to develop an implementation standard, and to work with agencies to discuss how crosswalking to the standard can be accomplished. Appendix B. Relation of USNVC to Land Cover ClassificationsTable B.1. Comparison of FAO LCCS Land Cover Types (based on structural domains) and National Land Cover Database (NLCD) types with that of NVC Level 1 (see Di Gregorio and Jansen 1996, USGS 2001).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rejecting Editorial Rejections: a Critique to Avoid Real Revisions
|
|
5–6 August 2006
|
|
Assymetric Competition in Marine Snails Littorine snails compete indirectly by altering the traits of a shared seaweed resource. Direct grazing by smooth periwinkles (Littorina obtusata; yellow snail on left) reduced seaweed palatability for conspecifics and congeners (common periwinkles, Littorina littorea; dark snail on right). In addition, previous grazing by smooth periwinkles reduced populations of common periwinkles on outplanted seaweeds. These interactions were asymmetric, with a specialist (smooth periwinkles) competitively superior to a generalist (common periwinkles). Littorine snails compete indirectly by altering the traits of a shared seaweed resource. Direct grazing by smooth periwinkles (Littorina obtusata; yellow snail on left) reduced seaweed palatability for conspecifics and congeners (common periwinkles, Littorina littorea; dark snail on right). In addition, previous grazing by smooth periwinkles reduced populations of common periwinkles on outplanted seaweeds. These interactions were asymmetric, with a specialist (smooth periwinkles) competitively superior to a generalist (common periwinkles). |
This photograph illustrates the article, “Asymmetric competition via induced resistance: specialist herbivores indirectly suppress generalist preference and populations” by Jeremy D. Long, Rebecca S. Hamilton, and Jocelyn L. Mitchell, tentatively scheduled to appear in Ecology 88(4), April 2007. |
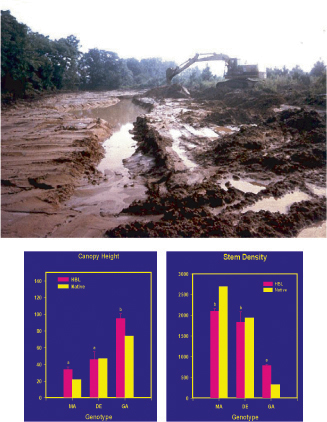
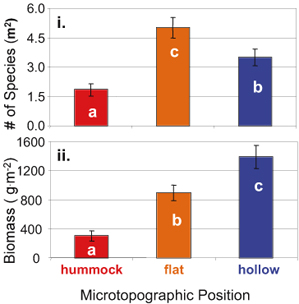
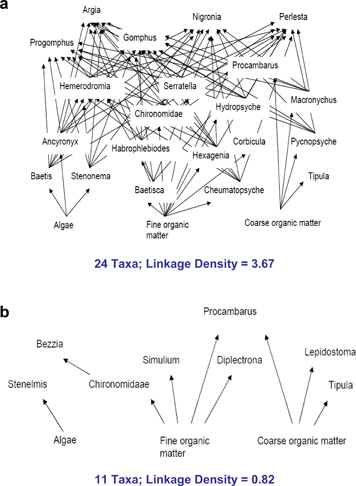
| The consequences of climate change for ecosystem structure and function remain largely unknown. A 20-year study of a high-elevation riparian system in Arizona shows substantial changes in both structure and function related to climate change. Abundances of dominant deciduous trees have declined dramatically over the 20 years, correlated with a decline in over-winter snowfall. Snowfall can affect over-winter presence of elk, whose browsing can significantly impact deciduous tree abundance. Seven species of birds were found to initiate earlier breeding associated with an increase in spring temperature across years, with minimal consequences for populations. Climate had much larger consequences for these seven bird species by affecting trophic levels below (plants) and above (predators) them. In particular, climate-related declines in deciduous vegetation led to decreased abundance of preferred bird habitat and increased nest predation rates. In addition, summer precipitation declined over time, and drier summers also were further associated with greater nest predation in all species. The net result was local extinction and severe population declines in some previously common bird species; one species increased strongly in abundance, and two species did not show clear population changes. Thus, climate can alter ecosystem structure and function through complex pathways that include direct and indirect effects on abundances and interactions of multiple trophic components.
|
 |
 |
Maple attempts to grow in the understory each year and here is the typical response, where any new shoots are browsed down by deer and elk. The density of understory maple stems has decreased by more than an order of magnitude over the 20 years of study. |
This aspen ramet is 6 years old and attempts to produce new shoots each year, but is browsed back so much that it is only 15 cm tall, whereas aspen released from browsing at this age are 5–8 m tall. The lack of aspen recruitment is leading to loss of aspen in the system; marked monitoring of 878 aspen over the past 10 years predicts it will be gone in about four more years. |
|
|
|
|
 |
|
|
These photographs illustrate the article, “Climate correlates of 20 years of trophic changes in a high-elevation riparian system,” by Thomas E. Martin, tentatively scheduled to appear in Ecology 88(2), February 2007. |
|
| Rocky intertidal mussels experience rapid fluctuations in aerial body temperature during low tide. Because of variability in tidal regimes and climatic regimes, geographic gradients in body temperature along the west coast of the United States may be much more complex than previous assumed. As a result, climate change may not lead to poleward range shifts, but instead may cause damage at a series of “hot spots.” All photos by Brian Helmuth, unless noted otherwise. |
|
 Rock intertidal zone at Santa Cruz Island, California. Photo credit: C. Blanchette. |
|
| Rocky intertidal zone at Boiler Bay, Oregon. The intertidal zone represents the interface between the terrestrial and marine environments. Thus animals and algae in these environments (such as Boiler Bay Oregon, shown here) may be among the first ecosystems to exhibit responses to global climate change. |
 |
 |
Strawberry Draw, Tatoosh Island, Washington State. The rocky intertidal zone has long served as an experimental laboratory for investigating the influence of climate on small-scale distribution patterns of organisms. |
| Mytilus. Results suggest that patterns of aerial body temperature (and in particular maximum temperature) may be complex, and thus patterns of mortality (shown here from Bodega Bay, California) may occur in unexpected locations. Photo credit: C. Harley. |
 |
 Intertidal zone at Tatoosh Island, Washington. |
|
 |
Sensors were deployed at mid-tidal heights in mussel beds at a series of sites in order to measure geographic patterns in aerial and aquatic body temperatures (Tatoosh Island, Washington). |
In their article, Helmuth et al. describe how latitudinal patterns of intertidal thermal stress may be more complex than anticipated. Using a series of biomimetic temperature loggers designed to mimic the thermal characteristics of mussels (Mytilus californianus), shown here, they measured temperature patterns at sites ranging from northern Washington to southern California. |
 |
 Intertidal zone at Tatoosh Island, Washington. |
|
| These photographs illustrate the article, “Mosaic patterns of thermal stress in the rocky intertidal zone: implications for climate change,” by Brian Helmuth, Bernardo R. Broitman, Carol A. Blanchette, Sarah Gilman, Patricia Halpin, Christopher D.G. Harley, Michael J. O’Donnell, Gretchen E. Hofmann, Bruce Menge, and Denise Strickland, which appeared in Ecological Monographs 76:461–480, November 2006. |
|
A long-term study, lead by Dr. Wayne Sousa (University of California, Berkeley), is investigating a variety of biotic and abiotic processes that may account for the structure and dynamics of mangrove forests on the Caribbean coast of central Panama. The study forests of Punta Galeta, near the Smithsonian Tropical Research Institute’s Galeta Marine Laboratory, contain three canopy tree species, which exhibit a pattern of shoreline zonation that is typical of the region. Seaward fringing stands are monopolized by Rhizophora mangle, low intertidal stands are a nearly even mixture of R. mangle and Laguncularia racemosa, and Avicennia germinans dominates more inland stands. L. racemosa often reappears in the canopy near the upland edge, and sometimes forms small monospecific stands along the mangrove-rain forest ecotone. Note: all photos by Wayne Sousa, with exception of Image 1, which is by Anand Varma. |
|
 Fringing stand of Rhizophora mangle at seaward end of tidal gradient. |
 Low intertidal mixed stand of Rhizophora mangle and Laguncularia racemosa. |
Upper intertidal stand of Avicennia germinans; seedlings on the forest floor are predominantly Avicennia. |
 |
Our forthcoming paper examines the roles of propagule dispersal and establishment in explaining species distributions along the tidal gradient. The propagules of mangroves are buoyant and dispersed by tidal currents and runoff following rain storms. We quantified dispersal patterns by monitoring the movements (directions and distances) of marked propagules released at different distances from the water’s edge. A separate experiment measured rates of seedling establishment at these same positions along the tidal gradient. Rabinowitz’ Tidal Sorting Hypothesis posits that zonation is the product of the (1) differential landward movement of propagules of different size by incoming tidal flow (i.e. tidal sorting) and (2) greater ability of larger propagules to establish in the deeper water characteristic of the low intertidal zone. Our observations were not consistent with this model of zonation, but revealed a different form of dispersal limitation that seems to strongly influence species’ vertical distributions and forest structure. |
|
 |
A small river running through a mixed stand of Rhizophora mangle and Lagunculariaracemosa. Mangrove propagules that disperse into a river channel can be transported longer distances. |
Dispersing propagules of Laguncularia racemosa trapped behind a log. |
 |
These photographs illustrate the article, “Supply-side ecology in mangroves: Do propagule dispersal and seedling establishment explain forest structure?” by Wayne P. Sousa, Peter G. Kennedy, Betsy J. Mitchell, and Benjamin M. Ordóñez L., tentatively scheduled to appear in Ecological Monographs 77(1):53–76, February 2007. |
|
|
Understanding Food Chains and Food Webs, 1700–1970 |
Some ecological ideas developed gradually and only gained coherence and details after they had become commonplace. The history of two interrelated ideas, food chains and food webs, is an example of a gradual, cumulative history. Here is a brief survey of these concepts from about 1700 to 1970 (Fig. 1). The earliest identified food chains seem to have concerned hyper-parasitism (Egerton 2005, 2006a), which students of insects discovered in the later 1600s. But it was entrepreneurial naturalist Richard Bradley (Egerton 2006b) who generalized the concept (Bradley 1718, part 3:60–61):
After reading this account, Rear-Admiral William Symonds told Darwin that he had seen at St. Paul crabs drag young birds from nests and eat them. Darwin added his information to this passage in the second edition (1845) of his book on the voyage of the Beagle (Edwards 1985:34).
In The Origin of Species (1859:73–74), Darwin reported the most famous example of a food chain in the scientific literature (Fig. 4).It is in chapter 3 on the “Struggle for existence,” and involves humble bees (called “bumble bees” in America) pollinating red clover; though some bees were eaten by field mice, the mice, in turn, were kept in check by domestic cats. Darwin speculated that if it were not for the cats, the mice would decimate the bees, and the clover would go unpollinated, since only humble bees pollinate clover. A later, unknown commentator extended this chain further (Milne and Milne 1966:6) by suggesting that old maids commonly kept cats, that clover-fed cattle were eaten by British seamen who protected the British Empire, and that if it were not for old maids, the British Empire would fall! In other words, Darwin’s food chain became a biological version of Englishman George Herbert’s well-known admonition (1640):
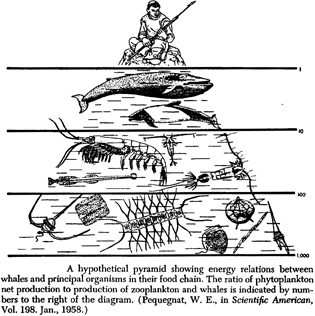
In America, this admonition is attributed to Ben Franklin, who borrowed it without acknowledgement for Poor Richard’s Almanac (1757). But getting back to Darwin’s food chain, in 1947 W. L. McAtee pointed out that Darwin’s food chain dynamic lacks full validity, since we now know that honey bees also pollinate red clover and that humble bees often appropriate mouse holes, so humble bees and mice have an ambiguous relationship. In Darwin’s defense, he heavily depended on H. W. Newman’s 1850–1851 study “On the habits of the Bombinatrices” (Darwin 1975:183). The next discussion of note for our purposes is from a remarkable German zoologist, Karl Semper. In 1877, he gave 12 lectures at the Lowell Institute in Boston, published simultaneously in English and German editions in 1881. The English title is Animal Life as Affected by the Natural Conditions of Existence. This book was the first detailed synthesis of animal ecology. In a discussion of the food of herbivores and carnivores (Semper 1881:51–52), he pointed out that when herbivores transform vegetation into flesh, there is a loss of mass due to oxidation of organic material, and that the same is true when carnivores transform the flesh of their prey into their own flesh. To illustrate this, he arbitrarily assumed a 10 to 1 ratio of food to flesh. One thousand units of plant food could only support 100 units of a herbivore, and those 100 units of herbivore could only support 10 units of a carnivore. Although his book has 106 illustrations, this generalized food chain was not illustrated. However W. E. Pequegnat’s diagram (Fig. 5) from Scientific American (1958:86) captures Semper’s concept, even to the point of using a 10 to 1 ratio. Semper wrote at a time when there was little quantified thinking in natural history. He had first trained as an engineer and then as a physiologist (Mayr 1975), and that background came to the fore in this discussion. Although his book was widely read, apparently no one carried this line of quantitative thinking any further in the 1880s or 1890s.
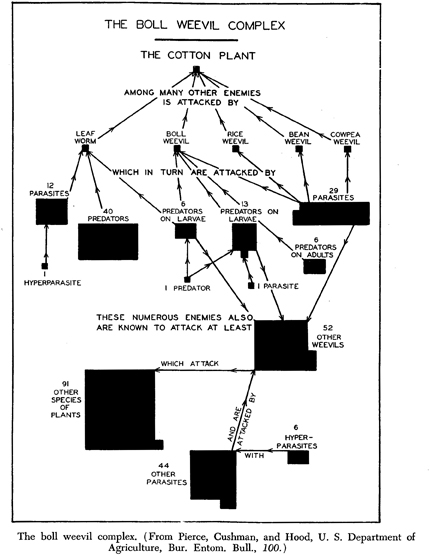
We are used to seeing food chains or webs diagrammed. The advantages are obvious: they provide a visual panorama of detailed information. The early history of such diagrams is elusive. The bibliography on food chains and webs that Allee, Emerson, Park, Park, and Schmidt compiled (1949:514) can assist in the search. However, they did not discover the earliest ones now known, published in 1880 by Lorenzo Camerano, which are reprinted in an English translation of his article (1994:377–378). Since Camerano’s two diagrams do not resemble any known from later zoologists, it seems likely that he did not have much, if any, influence on later diagrams. Joel Cohen (1994:353–355) suggests that Camerano was influenced by diagrams for other purposes in books by Darwin and by Hermann Helmholtz, though Camerano’s diagrams do not resemble theirs. Like Semper’s, Camerano’s food webs are generalized rather than specific. The earliest specific food web I have found (Fig. 6) is on “The boll weevil complex,” published in 1912 by Pierce, Cushman, and Hood in a USDA Bulletin. Their motive was to promote bowl weevil eradication—by encouraging its predators and parasites. Theirs may not have been the first specific diagram published, because others appeared about the same time in different biological specializations, where it is unlikely that the members of one specialization were reading the literature of other specializations.
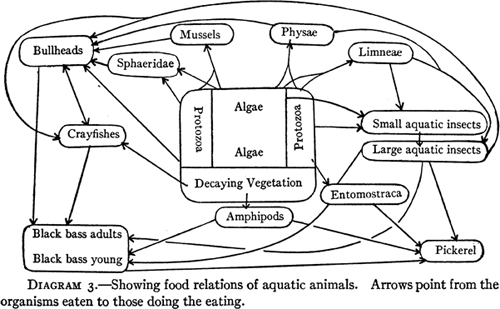
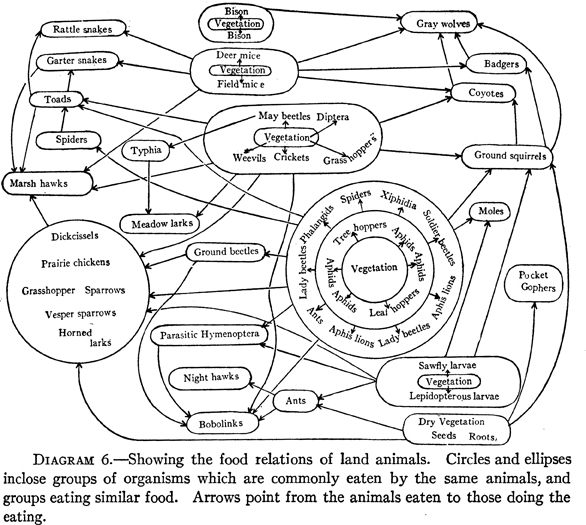
The following year, University of Illinois animal ecologist Victor E. Shelford (Fig. 7; photo, Croker 1991) published Animal Communities in Temperate America as Illustrated in the Chicago Region, which contained diagrams of both aquatic (Fig. 8) and land food webs (Fig. 9). There is no reason to suspect that he was influenced by the boll weevil diagram of 1912. Shelford used both of his diagrams to show how the community tends toward equilibrium, although the terrestrial community was more complex than the aquatic community, and consequently its equilibrium was more precarious. Shelford became a leading American animal ecologist (Croker 1991); his book was reprinted in 1937 and 1977.
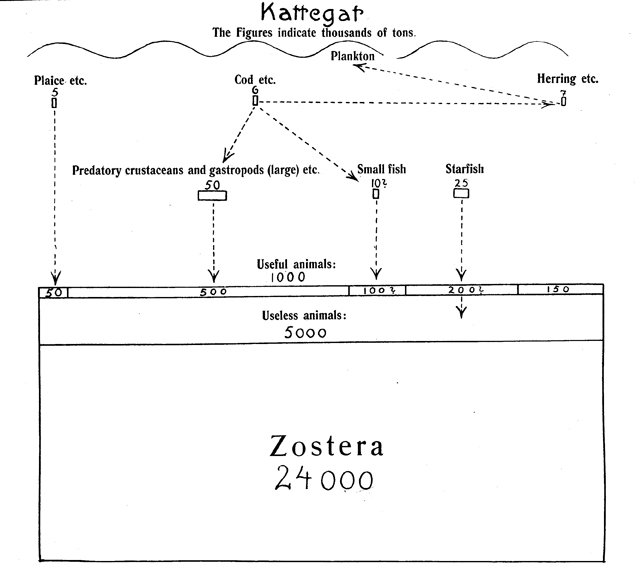
The earliest known food web diagram for a marine community was drawn by Danish fishery biologist Johannes Petersen (Fig. 10) in “A preliminary result of the investigations on the valuation of the sea”(1915). He studied the Kattegat region of shallow water between eastern Denmark and Sweden (Fig. 11), an area with maximum length of 150 miles and maximum width of 90 miles. Significantly, he attempted to establish the annual productivity for this region, and his diagram indicates the thousands of tons of each group of organisms, with both a number and a proportioned rectangle (Fig. 12). In the text he stated that the eel-grass (Zostera marina) figure of 24,000,000 tons represents only the amount produced in the summer, and that the annual production is twice that. Presumably, all the other figures are annual production and not just summer production. The tons of plaice and cod are the actual commercial catch of those fish from International Fishery Statistics for 1910, and that was possibly true also for the tons of herring given, though he did not say so. The numbers given for other animals seem to be estimates. Although he indicated on his diagram that herring fed on plankton, he thought plankton was much less important than Zostera as a foundation for this food web. He concluded that the Kattegat had a “very unfavourable proportion between producers and consumers”(Petersen 1915:32). What he meant by this seems to be indicated by the following sentence in which he stated that carp ponds have “even without artificial feeding, given a yield of fish per hectare several times greater than that of the Kattegat.” Petersen reproduced the same diagram with minor alterations in his final report, “The sea bottom and its production of fish-food”(1918:23).
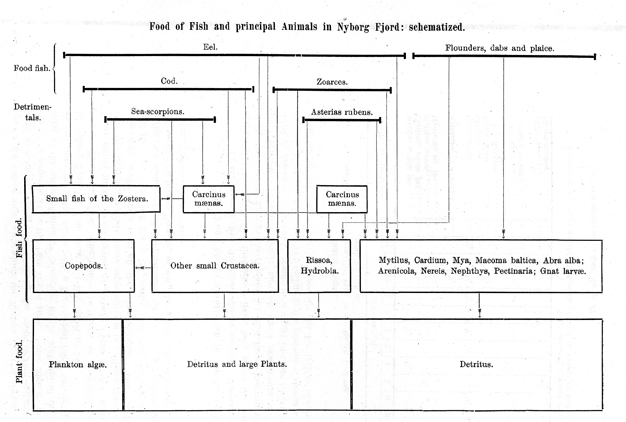
His colleague, H. Blegvad, also used rectangles in his diagram of “Food of fish and principal animals in Nyborg Fjord” (1916:24) (Fig. 13)but without attempting to represent precise quantities. However, he did give quantitative data in the text of his article, which provided some sense of the quantities of organisms involved at each level.
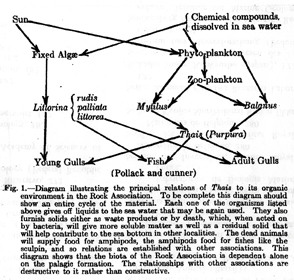
In the same year as Blegvad, the American zoologist Harold Sellers Colton published what Jonathan A. D. Fisher calls (2005:145) “possibly the first intertidal marine food web ever illustrated.” It is in Colton’s article on a carnivorous snail, Thais lapillus (now Nucella lapillus), and shows both which animals the snail eats and which animals eat the snail (Fig. 14). Colton did not indicate what inspired his diagram, but his brief bibliography does include Shelford’s book (1913). Fisher did not find references in the later relevant literature to Colton’s two articles on this snail (probably due in part to Colton’s leaving marine biology for archeology [Miller 1991]), so we do not know of any influence that his diagram exerted.
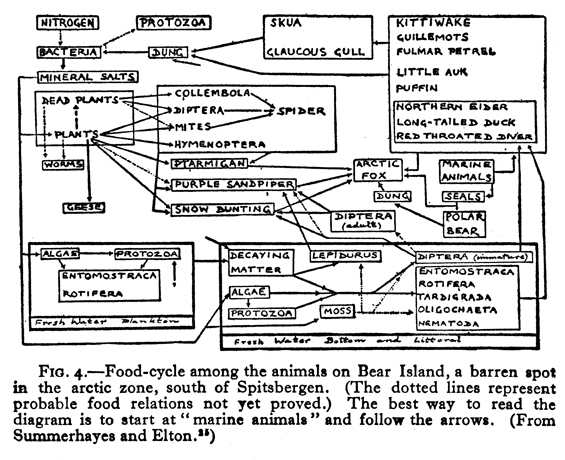
Charles Elton (Fig. 15) helped make such diagrams commonplace. He went on an Oxford University Arctic expedition in 1921 to Spitsbergen and took along Shelford’s book as a possible model for his own study (Elton 1966:33). However, Elton soon realized that the community he studied had a different dynamic than Shelford’s aquatic and terrestrial ones. Elton was impressed by the transfer of food from sea to land, which is reflected in his diagram (Fig. 16) published in 1923.
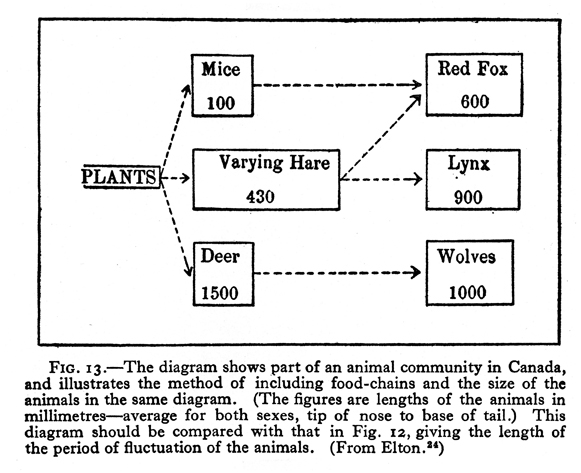
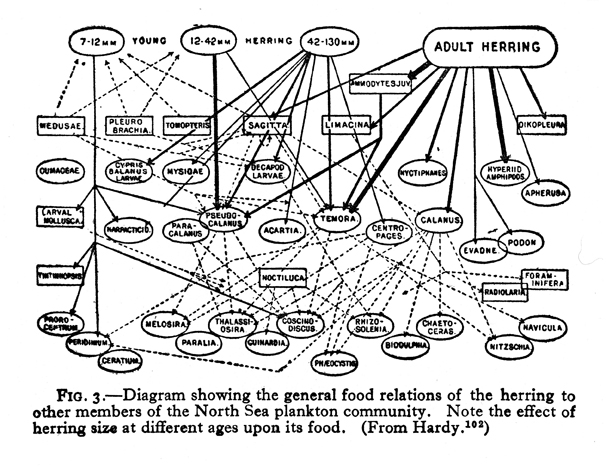
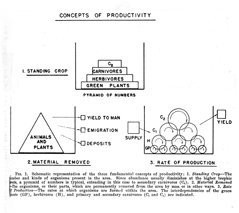
Although V. S. Summerhayes is listed as the senior author of their joint study, since he was a botanist, we can assume that Elton developed this diagram, in which plants are not emphasized. Two years later, in 1925, Elton published this much simpler Canadian food web (Fig. 17),which includes information on the lengths of animals. In 1924, English fishery biologist A. C. Hardy published a diagram (Fig. 18)on food consumed by herring at different stages of development. It bears no similarity to any diagrams previously shown, and it seems likely that he either was inspired by some unidentified example from the fisheries literature, or that he independently developed his diagram. Be that as it may, in 1927 Elton published his classic textbook, Animal Ecology, which reprinted and explained these last three diagrams by himself and Hardy. In that book Elton also introduced (1927:55) the terms “food chain” and “food cycle.” Widespread use of his book popularized the use of food web diagrams. In both Hardy’s diagram and in Elton’s for 1925, more information was conveyed than merely which animal ate which food. Hardy’s additional information was on the age of herring in relation to food, and Elton’s was on the size of the consumer in relation to food. Elton also popularized the idea of a food pyramid (1927:68–70), which concept had been implied by Semper.
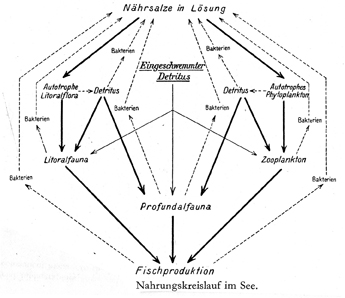
In 1926 Germany’s leading limnologist, August Thienemann (Fig. 19) published this unique food web of lakes (Fig. 20). His 50-page article on nutrient cycles in lakes introduced into limnology the terms “producers,” “consumers,” (though Petersen 1915 [quoted above] had used both terms in marine biology) and “reducers.”
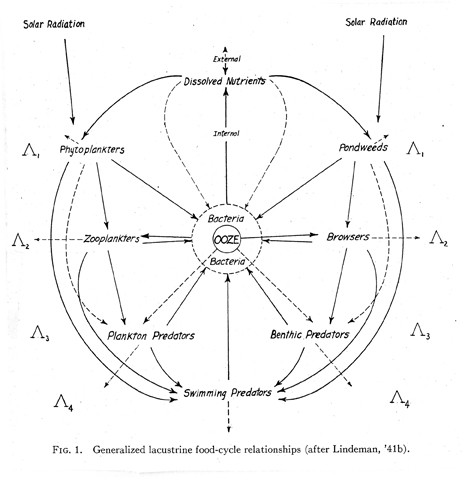
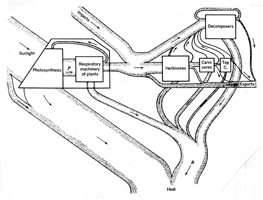
Thienemann’s 1926 paper and two of his other papers influenced an American postdoctoral student, Raymond Lindeman, who produced one of the most influential diagrams in the history of ecology (Fig. 21), though few if any ecologists have published similar diagrams.
It appeared in his posthumous paper, “The trophic–dynamic aspect of ecology” (1942). Like Thienemann’s diagram, Lindeman’s is a generalized food web, but both men had hard specific data backing up their concepts. In that respect theirs were similar to diagrams by Shelford, Elton, and Hardy, which illustrated specific food webs, and unlike Semper’s generalized food web, which was an educated guess. In the caption to his diagram Lindeman indicated that it was similar to one he had published the previous year. A comparison of his two diagrams indicates what he learned in his year at Yale University working under Evelyn Hutchinson (Cook 1977). The 1941 diagram is identical to the 1942 diagram except it lacks the symbols for trophic levels along the side. Lindeman (1942:159) used Thienemann’s terms “producers” and “consumers,” but suggested substituting the term “decomposers” for Thienemann’s term, “reducers,” to signify that the indicated process was not just chemical, but also biological.
In 1943, a year after Lindeman’s 1942 diagram appeared in the journal Ecology, Harvard marine ecologist George Clarke published this conventional food web (Fig. 22), but three years later, after he had studied Lindeman’s diagram and its explanation, Clarke published his diagram (Fig. 23) in Ecological Monographs, of a marine food web that emphasizes productivity and human removal of material. It also shows Clarke’s concern for the rate of production at each trophic level.
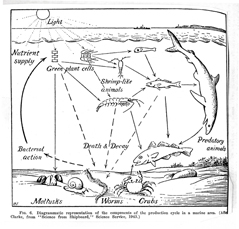
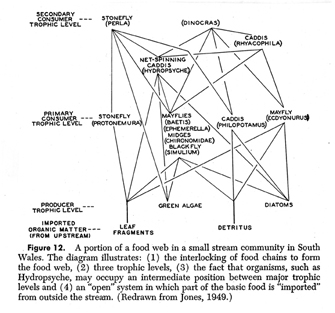
The Odum brothers, Eugene and Howard Thomas, carried Lindeman’s thinking further. The Atomic Energy Committee became interested in radiation ecology (Kwa 1989:48), and Eugene Odum (Fig. 24; photo, Craige 2001) developed a program at the University of Georgia to study food chains at the Savannah River Research Facility to trace radioactive pollution (Craige 2001). By injecting plant stems with radioactive phosphorus-32, he and his colleagues traced it up the food chain to leafhoppers, beetles, and spiders (Kwa 1989:58). About 1957 the programs at Oak Ridge and Savannah River converged, with both programs using radioactive tracers to measure the flow of materials up the food chain (Kwa 1989:66). In the second edition of Eugene Odum’s famous textbook, Fundamentals of Ecology (1959:47), there is a 1949 diagram of a food chain (Fig. 25). When I saw it, I assumed that Lindeman’s influence had flowed across the Atlantic in just a few years, but when I compared it with British ecologist Erichsen Jones’ own diagram, I discovered what Odum meant when he wrote that his diagram was “redrawn” from the one by Jones: Odum added the labels to the left of the diagram as a pedagogical aid.
Howard Thomas Odum (Fig. 26; photo from Katherine Ewel) received his graduate training under Hutchinson at Yale. (In 1954 he taught me freshman zoology at Duke.) In 1956, he produced a diagram (Fig. 27) of matter and energy flow, in steady-state flowing-water communities in Florida. At that point, the reader could still understand the diagram without special training. However, H. T. Odum continued developing his thinking along the lines of systems ecology and used symbols from electrical engineering. By 1971 he published esoteric diagrams (Fig. 28) that integrate humans into the biotic community. This was an important step towards founding several applied ecological sciences (Mitsch 1994, Hall 1995, Egerton 2007b). Other ecologists developed food chain and food web concepts in another direction. In 1948, D. E. Howell reported finding DDT in human fat, and by 1949 biologists were reporting that fish feeding on insects killed by DDT were also being killed (Hoffmann and Surber 1949, Langford 1949).
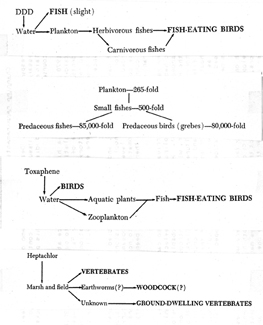
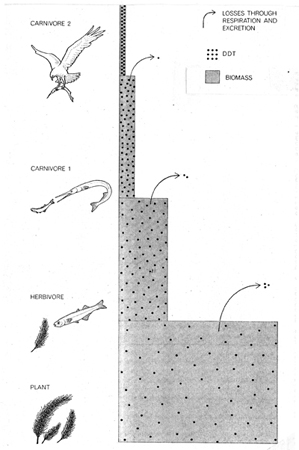
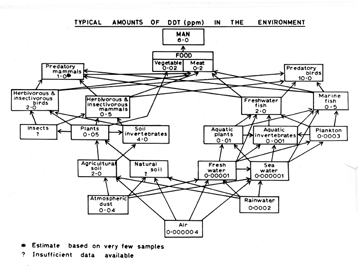
Rachel Carson (Fig. 29) publicized the discovery of insecticides traveling up the food chain in ever-increasing concentrations in her best-selling book, Silent Spring (1962:110–111), as did Robert Rudd in his less-read book, Pesticides and the Living Landscape (1964). Carson did not provide diagrams, and the ones Rudd used were quite simple. Here are four (Fig. 30) of the seven diagrams in his book. DDT was the most notorious insecticide, and in 1967 George Woodwell published a diagram (Fig. 31) in Scientific American showing increased concentrations of DDT as it progressed up the food chain. By 1970, Clive Edwards constructed a much more detailed food web (Fig. 32), showing DDT pathway and concentrations from the time of spraying DDT into the air, all the way up the food chain until it became concentrated in predatory birds, mammals, and humans.
From simple narratives around 1700, food chain and food web concepts have been developed into progressively more sophisticated vehicles for conveying ecological ideas (Polis et al. 2004, de Ruiter et al. 2005). Lorenzo Camerano’s two 1880 diagrams of food webs had no known influence, but after the visual stimulus of diagrams became established in the early 1900s, many ecologists found creative ways to express visually their discoveries concerning food chains and webs. Literature cited Allee, W. C., A. E. Emerson, O. Park, T. Park, and K. P Schmidt.1949. Principles of animal ecology. W. B. Saunders, Philadelphia, Pennsylvania, USA. Blegvad, H. 1916. On the food of fish in the Danish waters within the Skaw. Report of The Danish Biological Station 24:17–72 + 6 lists. Bradley, R. 1718. New improvements of planting and gardening, both philosophical and practical. Edition 2, Part 3. [Edition 1, 1717–1718.] W. Mears, London, UK. Camerano, L. 1880. Dell’ equilibrio dei viventi mercè la reciproca distruzione. Atti della Reale Accademia delle Scienze du Torino 15:393–414 + 4 plates. Camerano, L. 1994. On the equilibrium of living beings by means of reciprocal destruction. Claudia M. Jacobi, translator. J. E. Cohen, editor. Pages 360–380 in S. A. Levin, editor. Frontiers in mathematical biology. Springer-Verlag, Berlin, Germany. Carson, R. 1962. Silent spring. Houghton Mifflin, Boston, Massachusetts, USA Clarke, G. L. 1946. Dynamics of production in aquatic populations. Ecological Monographs 16:321–335. Cohen, J. E. 1994. Lorenzo Camerano’s contribution to early food web theory. Pages 351–359 in S. A. Levin, editor. Frontiers in mathematical biology. Springer-Verlag, Berlin, Germany. Cook, R. E. 1977. Raymond Lindeman and the trophic–dynamic concept in ecology. Science 198:22–26. Craige, B. J. 2001. Eugene Odum: ecosystem ecologist and environmentalist. University of Georgia Press, Athens, Georgia, USA. Croker, R. A. 1991. Pioneer ecologist: the life and work of Victor Ernest Shelford, 1877–1968. Smithsonian Institution Press, Washington, D.C., USA. Darwin, C. R. 1839. Journal of researches into the geology and natural history of the various countries visited by H.M.S. Beagle, under the command of Captain Fitzroy, RN from 1832 to 1836. Henry Colburn, London, UK. Darwin, C. R. 1859. On the origin of species by means of natural selection. John Murray, London, UK. Darwin, C. R. 1975. Natural selection: being the second part of his big species book written from 1856 to 1858. R.C. Stauffer, editor. Cambridge University Press, Cambridge, UK. De Ruiter, P. C., V. Wolters, and J. C. Moore, editors. 2005. Dynamic food webs: multispecies assemblages, ecosystem development, and environmental change. Academic Press, Boston, Massachusetts, USA. Edwards, A. J. 1985. Saint Paul’s Rocks: a bibliographical review of the natural history of a mid-Atlantic island. Archives of Natural History 12:31–49. Edwards, C. A. 1970. Persistent pesticides in the environment. CRC Press, Cleveland, Ohio, USA. Egerton, F. N. 1973. Changing concepts of the balance of nature. Quarterly Review of Biology 48:322–350. Egerton, F. N. 2005. A history of the ecological sciences, part 17: invertebrate zoology and parasitology during the 1600s. ESA Bulletin 86:133–144. Egerton, F. N. 2006a. A history of the ecological sciences, part 19: Leeuwenhoek’s microscopic natural history. ESA Bulletin 87:47–58. Egerton, F. N. 2006b. A history of the ecological sciences, part 20: Richard Bradley, entrepreneurial naturalist. ESA Bulletin 87:117–127. Egerton, F. N. 2007a. Carl Linnaeus and the economy of nature. ESA Bulletin 88, in press. Egerton, F. N. 2007b. History of ecology. In S. E. Jørgensen, editor. Encyclopedia of ecology. Elsevier, Oxford, UK. Elliott, A. M. 1957. Zoology. Edition 2. [Edition 1, 1952.] Appleton-Century-Crofts, New York, New York, USA. Elton, C. S. 1927. Animal ecology. Sidgwick and Jackson, London, UK. Elton, C. S. 1966. The pattern of animal communities. Methuen, London, UK. Fisher, J. A. D. 2005. Exploring ecology’s attic: overlooked ideas on intertidal food webs. ESA Bulletin 86:145–151. Hall, C. A. S., editor. Maximum power: the ideas and applications of H. T. Odum. University Press of Colorado, Niwot, Colorado, USA. Hegner, R. 1938. Big fleas have little fleas, or who’s who among the protozoa. Williams and Wilkins, Baltimore, Maryland, USA. Hoffmann, C. H., and E. W. Surber. 1949a. Effects of feeding DDT-sprayed insects to freshwater fish. U.S. Fish and Wildlife Service, Special Science Report (Fish), number 4, 9 pages. Hoffmann, C. H., and E. W. Surber. 1949b. Effects of an aerial application of DDT on fish and fish-food organisms in two Pennsylvania watersheds. Progressive Fish-Culture 11:203–211. Howell, D. E. 1948. A case of DDT storage in human fat. Oklahoma Academy of Science Proceedings 29:29–31. Jones, J. R. E. 1949. A further ecological study of a calcareous stream in the “Black Mountain” district of South Wales. Journal of Animal Ecology 18:142–159. Kwa, C. L. 1989. Mimicking nature: the development of systems ecology in the United States, 1950–1975. University of Amsterdam, Amsterdam, The Netherlands. Langford, R. R. 1949. The effects of DDT on freshwater fishes. Pages 19–38 in Forest spraying and some effects of DDT. Canada Department of Lands and Forest Biological Bulletin Number 2. Lindeman, R. L. 1941. Seasonal food-cycle dynamics in a senescent lake. American Midland Naturalist 26:636–673. Lindeman, R. L. 1942. The trophic–dynamic aspect of ecology. Ecology 23:399–418. Linnaeus, C. 1749. Specimen academicum de oeconomia naturae. I. J. Biberg, respondent. Upsala, Sweden. Linnaeus, C. 1775. Miscellaneous tracts relating to natural history, husbandry, and physick. B. J. Stillingfleet, translator. Third edition. J. Dodsley, Baker and Leigh, London, UK. Linnaeus, C. 1977. Miscellaneous tracts relating to natural history, husbandry, and physick. B. J. Stillingfleet, translator. Arno Press, New York, New York, USA. McAtee, W. L. 1947. The cats-to-clover chain. Scientific Monthly 65:241–242. Mayr, E. 1975. Carl Gottfried Semper (1832–93). Dictionary of Scientific Biography 12:299. Miller, J. H. 1991. The life of Harold Sellers Colton: a Philadelphia brahmin in Flagstaff. Navajo Community College Press, Tsaile, Arizona, USA. Milne, L. J. and M. Milne. 1966. The balance of nature. Knopf, New York, New York, USA. Mitsch, W. J., editor. 1994. Energy flow in a pulsing system: Howard T. Odum. Ecological Engineering 3:77–105. Newman, H. W. 1850–1851. On the habits of the Bombinatrices. Entomological Society of London Transactions, Proceedings, New Series 1:86–92, 109–112, 117–118. Odum, E. P., in collaboration with H.T. Odum. 1959. Fundamentals of ecology. W. B. Saunders, Philadelphia, Pennsylvania, USA. Odum, H. T. 1956. Primary production in flowing waters. Limnology and Oceanography 1:102–117. Odum, H. T. 1971. Environment, power, and society. Wiley-Interscience, New York, New York, USA. Pequegnat, W. E. 1958 (January). Whales, plankton and man. Scientific American 198(1): 84–90. Petersen, C. G. J. 1915. A preliminary result of the investigations on the valuation of the sea. Report of the Danish Biological Station 23:29–33. Petersen, C. G. J. 1918. The sea bottom and its production of fish-food. Report of the Danish Biological Station 25:1–66 + 10 plates + map. Pierce, W. D., R. A. Cushman, and C. E. Hood. 1912. The insect enemies of the cotton boll weevil. U.S. Department of Agriculture, Bureau of Entomology Bulletin 100:1– 99. Polis, G. A., M. E. Power, and G. R. Huxel. 2004. Food webs at the landscape level. 2004. University of Chicago Press, Chicago, Illinois, USA. Semper, K. G. 1881. Animal life as affected by the natural conditions of existence. Appleton, New York, New York, USA. Reprint 1977. Arno Press, New York, USA. Shelford, V. E. 1913. Animal communities in temperate America as illustrated in the Chicago region. University of Chicago Press, Chicago, Illinois, USA. Reprint 1937 with bibliographic addenda. Reprint 1977. Arno Press, New York, New York, USA. Summerhayes, S. V., and C. S. Elton. 1923. Contribution to the ecology of Spitsbergen and Bear Island. Journal of Ecology 11:214–268. Swift, J. 1733. On poetry: a rhapsody. J. Huggonson, London, UK. Thienemann, A. F. 1926. Der Nahrungskreislauf im Wasser. Verhandlungen der deutschen zoologischen Gesellschaft 31:29–79. Woodwell, G. M. 1967 (March). Toxic substances and ecological cycles. Scientific American 216(3):24–31. Acknowledgments This is a revised version of a talk given at the ESA Annual Meeting in August 2006 in Memphis, Tennessee. For comments preceding the talk, I thank Robert P. McIntosh, Professor Emeritus of Biology, University of Notre Dame (now in Florida). For several references used in the revision, I thank Jonathan A. D. Fisher, Department of Biology, University of Pennsylvania, Philadelphia. Frank N. Egerton |
||||||||||||||||||||||||||||||
Commentary |
|
AGU Editorial: Advocacy and Integrity
|
|
Advocacy and Integrity |
|
Commentary |
A History of the Ecological Sciences, Part 23: Linnaeus and the Economy of Nature |
| Carl Linnaeus (1707–1778) was a leading naturalist of the 1700s (Lindroth 1973, 1983, Morton 1981:259–276, 281–285, Goerke 1993, Broberg 2000, Spary 2002). All ecologists know he founded modern nomenclature and systematics (Larson 1971, Stafleu 1971, Mayr 1982:171–180, Eriksson 1983), but he is less well known for inventing an ecological science he called the economy of nature. He explained it in 1749, but the overly broad science of natural history, which he had pursued since childhood, was already ecological in outlook and content. In 1749 he generalized and formalized what he had been recording specifically and informally. A series of 186 essays, largely by Linnaeus, were defended by his students as dissertations for their doctoral degrees (Jackson 1913, Ramsbottom 1959:151–153, Smit 1989:118–119, Kiger et al.1999:231), and one of these was Specimen academicum de oeconomia naturae (1749), defended by Isaac J. Biberg. Linnaeus republished these dissertations in 10 volumes entitled Amoenitates Academica (Academic Pleasures, 1749–1790), though the last two volumes appeared posthumously. The Amoenitates Academica has been reprinted several times, and 19 dissertations are translated into English (Linnaeus 1775, 1781, 1977a, b). There is a helpful Index to Scientific Names of Organisms cited in Linnaean Dissertations (Kiger et al. 1999), with a guide to collected editions. Linnaeus’ earlier natural history observations are recorded in travel books and other writings. |  Fig. 1. Linnaeus in his Lapland clothes (or costume). Drawn in Holland by Martin Hoffman, 1737. |
|||||||||||||||||||||||||||||||
All of his travel books and the dissertations are listed in B. H. Soulsby’s catalogue of Linnaeus’ works (1933:23–26, 99–151). Florence Caddy (1886–1887) provides two good maps on Linnaeus’ travels, though the caption to the one at the end of volume I is misdated 1735–1738 (read 1732–1738). Wilfrid Blunt (1971) includes maps and summaries of the trips in his biography of Linnaeus. Linnaeus’ travel books show his broad interest in plants, animals, geology (Merriam 2004), and economic uses of natural history (Linnaeus 1766, 1781:1–67, 1977, Koerner 1999, Müller-Wille 2003, Rausing 2003). David Black selected natural history extracts from Linnaeus’ books on the 1732 and 1741 trips, which he published with a map and modern illustrations by Stephen Lee (Linnaeus 1979). Linnaeus’ first expedition was undertaken in 1732, begun on 12 May, his 25th birthday, and lasted until 10 October. He traveled north to Lapland and then west to the Norwegian coast. On the return journey he traveled in Finland down the eastern side of the Gulf of Bothinia to Åbo and then crossed to Stockholm. It was his longest journey—he estimated 633 Swedish miles or about 3800 English miles (1811, II:270, 1971)—and the subject of his longest travel book. It was also the one travel book that he illustrated. A historian of Swedish botany judged this trip “the most productive exploratory expedition ever undertaken in Sweden”(Fries 1950:18). It was sponsored by the Royal Academy of Sciences at Uppsala, which declined to publish his manuscript, and an English translation was published (1811) long before the Swedish version (1913). Linnaeus’ most recent biographer, who reads Swedish, judges some of his behavior and writings on this trip rather harshly (Koerner 1999:59–65). She says he doubled the actual distance he traveled in his report (her figure is 4500 miles) because he was to be paid per mile, that he drew a map indicating travel to places he had not visited, and that he later claimed to have stayed in Lapland much longer than he had. While I cannot check all her claims, this statement is doubly wrong: “He never passed the sixtieth degree north Latitude, which marks the Arctic Circle” (Koerner 1999:61). The Arctic Circle is actually at 66°30’, and he did cross it. He visited Jokkmokk (29 June) just north of that line, and more than half a dozen other places north of Jokkmokk (see map in Blunt 1971:41). She does not accuse him of inventing any of his natural history observations. When he visited the cataract of the Elf-Carleby River on 13 May, he described the salmon fishery below the cataract, the foam and spray that the cataract generated, and surrounding plants. But he did not merely describe; he also pondered how species lived (Linnaeus 1811, I:13, 1971):
Linnaeus’s illustrations did not always represent what he discussed in most detail. For example, he made an excellent drawing of a crane fly but only recorded that he collected it at Umeå on 9 June.
Remarkably, he shot a hawk owl from his horse, “going on at a good rate” (Linnaeus 1811, I:204, 1971), at 12:15 am. Regrettably, it was too damaged by the shot to be stuffed, but at least he drew its picture.
For two insects collected at Lulea on 21 June, he provided both illustrations and discussion (Linnaeus 1811, I:233, 1971)
The scientific names of species he discussed or illustrated were added by the editor, James E. Smith, from Linnaeus’s Flora Lapponica (1737) and Fauna Svecica (1746). He also discussed at length a rhododendron he drew on 8 July in the Lapland Alps (Fig. 5), but only to describe it and to evaluate whether it belonged in the azalea genus (Linnaeus 1811, I:299–301, 1971). On the Norwegian coast at Torfjorden he went fishing in a boat and caught with hook and line “plenty” of Sey-fish (Gadus virens), which he drew (Fig. 6). He found remora sticking to some of these 10-inch fish.
He also observed, collected, and drew four different kinds of medusa (jellyfish), but made no observations on their behavior or food (Linnaeus 1811, I:336–339, 1971). Two later Linnaean dissertations were on marine subjects: Noctiluca Marina (1752) on minute phosphorescent “insects,” and Natura Pelagi (1757) on fish, turtles, and cetaceans (Smit 1979:120–123). Reindeer were important draft and milk animals to Laplanders, and Linnaeus discussed them repeatedly in Lachesis Lapponica (1811), but only illustrated bridle, harness, and antlers (Linnaeus 1811, I:103–110, 135, 1971). He illustrated reindeer themselves in the frontispiece of Flora Lapponica, 1737). Their antlers were beginning to sprout in June, initially covered by soft skin which was often bloody from mosquito bites.
Females have smaller antlers than males. Squirrels gnawed antlers from previous years (Linnaeus 1811, I:127–128, 1971). Linnaeus’ discussion of what reindeer eat is interesting (Linnaeus 1811, I:161–162, 1971):
But he also reported that they ate nothing in hot weather, when mosquitoes were very troublesome (Linnaeus 1811, I:308, 1971). Later, he commented that the Lapps were negligent not to gather Lichen rangiferinus and horsetail (Equisetum fluviatile) in summer for winter fodder (Linnaeus 1811, II:107–108, 1971). Females give birth in May and fawns grow simple antlers their first year (Linnaeus 1811, I:313, 1971). In warm weather reindeer are tormented by the bites of gadflies (Oestrus tarandi), which leave so many scars that one author mistakenly thought they were caused by smallpox. (For more details on this fly and reindeer, see Linnaeus 1739, 1746b; part of the latter is translated by Susan Novikoff in Usinger 1964:5–6.) One insect, “probably a species of Tabanus” (Linnaeus 1811, I:280–281, 1971) bores into reindeer and lays its eggs under the skin, and the young leave by the same hole. The Lapps squeeze out the larva from their pustules to lessen the reindeer’s pain. Another fly (Oestrus nasalis) lays eggs in reindeer nostrils (Linnaeus 1811, II:45, 1971). Reindeer also suffer from an epidemic disease that Laplanders called Pekke Kattiata that could be fatal (Linnaeus 1811, II:39–40, 1971). These observations were also included in a 1754 dissertation, Cervus Rheno, defended for a doctorate degree by Charles F. Hoffberg, and is translated into English (Linnaeus 1781:167–214, 1977). On 17 July 1732, Linnaeus had a chance to see lemmings, which he described, and said they ate grass and reindeer moss. They lived mainly in the Scandinavian alps, but (Linnaeus 1811, II:19, 1971):
(The parenthetic question about burrow depth was inserted by the book’s editor.) Later in the book, Linnaeus raised his estimate of their numbers from thousands to millions and admitted that “nobody knows what becomes of them” (1811, II:82–83). In a still later article (Linnaeus 1740; partly translated in Blunt 1971:60), he rejected the belief that lemmings fall from clouds.
Without publishing his travel journal, Linnaeus still publicized his achievements, and the governor of Dalecarlia province offered to fund a survey of that province. Linnaeus agreed, and seven medical students gained permission to come along at their own expense. They first traveled to Falun, the provincial capital (where Linnaeus met his future wife), and then departed on their expedition on 3 July 3 1734, taking along the governor’s two sons. Linnaeus was an organizing genius, and he delegated specialized tasks to each student: geography; climate and soils; stones, minerals and fossils; plants; animals; economics; and logistics. Every night each student added his report to whatever Linnaeus wrote. The last entries were dated 17 August, and when they returned to Falun, Linnaeus gave their Iter Dalecarlium to the governor. It was never published, but some account of the trip appeared in a Hamburg newspaper, and Linnaeus used some of their notes in later publications (Blunt 1971:76–79, Caddy 1886–1887, I:213–249). Linnaeus did publish observations from subsequent field trips, and the book on his trip to Öland and Gotland in 1741 is also translated into English. The government (Swedish Estates of the Realm) asked him to make an economic survey, including natural history, of these islands. Accompanied by six young men, he departed from Stockholm on 15 May. It was quite cold, and Linnaeus suggested that “Spring should be measured according to climate and temperature rather than by the calendar”(Linnaeus 1973:23), and he then gave what we call phenological observations on the progress of the leaves and flowers or buds of several trees and herbs. Back in 1737 he had publicized a thermometer in the frontispiece to his Hortus Cliffortianus.
The thermometer was probably one he had obtained during three months spent in England, and he may have suggested to his friend Anders Celsius (1701–1744) that he reverse the scale he had developed, having boiling water at zero and freezing at 100 (Nordenmark 1935), because on 30 October 1758 Linnaeus wrote to a Montpellier botanist, Boissier de la Croix de Sauvages (English translation in Middleton 1966:100):
Two subsequent dissertations were phenological: Vernatio Arborum (1753) and Calendarium Florae (1754), and are translated into English (Linnaeus 1775:133–158, 233–286, 1977). When our explorers reached the copper smelter at Adelfors on 23 May 1741, Linnaeus noticed that the junipers looked like “trimmed cypresses”(Linnaeus 1973:34), which he attributed to smoke from the blast furnaces. Workers and residents at Adelfors complained about the air pollution. They reached Öland on 1 June, and Linnaeus made an inventory of its plants and animals. A gamekeeper told him the time of mating and the gestation periods of red and fallow deer, wild boar, and bear, which he recorded (Linnaeus 1973:48). He examined the nest of a Rook (Corvus frugilegus) containing three nestlings and numerous mites (Simulium reptans) bloated with nestlings’ blood. He counted annual rings of an oak stump and found it was 260 years old. Some rings were wider than others, which he thought was due to different severities of winters (Linnaeus 1973:58). Although modern botanists correlate annual ring width with summer moisture, this was a beginning of paleoclimatology. He knew that Francesco Redi had described 30 kinds of bird mites, which inspired Linnaeus (1973:69) to describe oystercatcher mites (Saemundssonia haematopi) and avocet mites (Vanellus vanellus). Along the seashore, he discovered that all plant species had succulent leaves, but that the majority of them growing elsewhere had ordinary dry leaves (Linnaeus 1973:72). Potentilla anserine grew on the sand and Senecio vulgaris on rotting seaweed. Cinnabar moths (Hipocrita jacobeae) were numerous on shore, and their larva ate the Senecio (Linnaeus 1973:86). He found that other plant species also had their own particular caterpillars, which he described and named, probably assisted by the entomologist Charles de Geer (Landin 1972), whom he visited at Medevi on 23 August (Linnaeus 1973:89, 199). Near the Lummelunda church he studied a marsh in which the sedge Cladium mariscus grew. This species had not previously been reported in Sweden; he emphasized the facts that cattle ate it in early spring and that it made good thatch for roofs. Since he learned that it grew in a former lake, he suggested that it be planted in Sweden’s many “sterile and useless bogs” that could not be drained (Linnaeus 1973:113). Beyond Stenkyrka, he found under stones in water a white oval leech (Hirundo [Nephelis] octoculata) that could also be found in the stomachs of small fishes, and he thought that the liver worms of sheep were probably the “spawn” of this leech, which the sheep swallowed when grazing in marshy places (Linnaeus 1973:118–119). After transcribing runic inscriptions in the Hangvar churchyard on 27 June, he commented that a white lichen (Kecabira cakcarea) grew on the limestone tombstones but not on granite ones (Linnaeus 1973: 119). They reached Fårö Island, just north of Gotland and much smaller (see map, Linnaeus 1973:facing page 109), on 28 June. Its inhabitants hunted seals but not porpoises. They also ate eider and their eggs, but Linnaeus thought that “The time will probably come when the excellent down of these birds will save them from being shot” (Linnaeus 1973:126), but he did not explain how to collect it (possibly from their nests). He described in some detail the growth of “sandhafre” (Ammophila arenaria) on the sand dunes, and explained how it stabilized the dunes. He also found ant lions on the dunes that were “far more multicoulored than on Öland” (Linnaeus 1973:130). He referred the reader to Réaumur’s memoir on ant lions for details.
Five years later, from 12 June to 11 August 1746, Linnaeus traveled through West Gothland and published his findings in Wästgöte-Resa (1747). Caddy (1886–1887, II:165–206) summarized this book, turning it into a Linnaean travelogue (she followed his route). Among the translated extracts quoted by Blunt (1971:163) is this generalization:
These thoughts were not especially original (Isaiah 40:6 “All flesh is grass.”), but they are of interest as a prelude to the 1749 dissertation on the economy of nature. Meanwhile, in 1744, the dissertation Oratio de telluris habitabilis incremento (On the increase of the habitable earth), defended by Johann Westmann, offered a novel geological theory (Frängsmyr 1983) and explanation of how the world had become populated with species (Linnaeus 1781:71–127, 1977b): (1) God created one pair of each sexual species and one individual of each hermaphroditic species; (2) since Adam named all species, the Garden of Eden must have been a mountain island; (3) each species increased in numbers every generation; (4) as they increased, they enlarged the geographical area they inhabited; and (5) the habitable land increased as the numbers of organisms increased. To support this argument, Linnaeus had to demonstrate the potential of all species to increase their populations. He listed the numbers of seeds reported for different flowers: Helenium 3000, Zea 2000, Helianthus 4000, Papaver 3200, and Nicotina 40,320. He then calculated correctly that an annual plant that only produced two seeds per year, if preserved from animals and accidents, would have 1,048,576 descendants in 20 years. That dissertation was only one of several publications that entitle Linnaeus to be called the founder of plant geography (Hofsten 1916:243–247, Browne 1983:16–23). Du Rietz (1957a) summarized his contributions to alpine phytogeography, paludology (on which see also Du Rietz 1957b), indicator plants, plant succession, limnology, and forest geography. A fundamental difference between Linnaeus’ conception of an ecological science and ours is that in his, biotic interrelationships were designed by God to work harmoniously and permanently and for the benefit of humanity (Hofsten 1957:90–102), whereas in ours, interrelationships evolve and can lead to extinction of species. His conception was part of a general outlook in science: for example, in astronomy, celestial bodies were unchanging in substance and orbits; and in geology, ongoing changes in the landscape were considered minor compared to the changes caused by God in the Flood of Noah. Scientists’ study of a “static” universe gradually revealed that it is not static. This even happened to Linnaeus. In his Systema Naturae (1735, 1964) he confidently claimed that all species had been created by God at the beginning and no new ones had since appeared. However, the discovery of Peloria in 1741—so similar to Linaria, yet an apparently different species—shook his belief in the constancy of species. He eventually suspected that God had created only a few species, which later hybridized to form the great variety now seen (Hagberg 1952:196–205, Hofsten 1957:65–86, Larson 1971:94–121, Bowler 1989:64–68). On 18 August 1764, he explained this idea in a letter to Johannes Burmann (in Nicolas 1963:53).
Linnaeus’ term “oeconomia naturae” (1749) is rather similar to the contemporary term for animal physiology, “animal economy,” which involved studying how the parts contributed to the functioning of the whole. He may have implied an analogy between organs in an animal and species in a biotic community (Linnaeus 1775:39, 1977a):
Having a passion for system, Linnaeus approached the economy of nature systematically. For each of the three kingdoms—stones (and soils), plants, and animals—he discussed a cycle of propagation, preservation, and destruction. Surveying different kinds of stones under “Propagation,” he suggested that one or more kinds had organic origins (Linnaeus 1775:51, 1977):
Under “Preservation,” he speculated, inaccurately, on how stones are generated and augmented by water, but under “Destruction” he was more accurate in describing the actions of weather and water in the gradual erosion of rocks. He also noted that certain animals also helped erode some kinds (Linnaeus 1775:57, 1977):
God allegedly designed living beings to both survive and regulate each other (Linnaeus 1775:40, 1977):
This explicit statement was an important contribution to the balance of nature concept, though Linnaeus did not name it (Egerton 1973:335–337). Under plant propagation, he discussed sexual reproduction, then seed dissemination (Linnaeus 1775:64–65, 1977):
Under “Preservation,” he claimed that God had decreed (Linnaeus 1775:67–68, 1977): “that the whole earth should be covered with plants, and that no place should be void, none barren.” He had heard of deserts but had never seen one, so he confidently asserted that they have their own unique trees and herbs (which they do, but there is still bare ground). All environments—alpine, grassland, marshes, aquatic, deserts—have characteristic species, and he discussed examples. The graesmasken moth inhibits the spread of grass, leaving room for other plants. However, plants die, and their destruction is also part of God’s plan. Black mould, which nourishes new plants, comes from dead plants, and that cycle really begins with the liverworts that grow on bare rocks; when they die, they leave mould for mosses, and as mosses die, they leave mould for herbs and shrubs. This dissertation contains one of the earliest descriptions of plant succession (which Clements, 1916:10, credited to Biberg, the defendant). Insects contribute to the death of plants by eating parts, which make them vulnerable to other hazards (Linnaeus 1775:76–80, 1977). Under animal propagation, Linnaeus surveyed all the known reproductive habits of different species, and although he rejected spontaneous generation, he admitted that (Linnaeus 1775:89),
He pointed out that smaller animals tend to produce more offspring than larger ones: mites can increase to a thousand in a few days, but elephants only produce one offspring in two years. However, some hawks are smaller than the poultry they eat, and he acknowledged that hawks layer fewer eggs, without attempting to explain why. He calculated that two pigeons breeding nine times a year could produce 14,672 young in four years, but his translator pointed out that Linnaeus had mistakenly added in the original pair to reach this figure (Linnaeus 1775:90, 1977); however, the numbers 6 and 7 were accidentally transposed in the English edition; Linnaeus’ figure should have been 14,760 (Egerton 1967:174). In Politia Naturae (Latin, 1760, cited from the English translation, 1781:162, 1977b), he added that long-lived animals propagate slowly. Under animal preservation, he discussed which species care for their young and which do not. Among polygamous species, “males scarcely take any care of the young” (Linnaeus 1775:93, 1977a), and cuckoos lay their eggs in the nests of wagtails and hedge-sparrows. Because of the great diversity of species, God assigned each one certain places to live and certain foods to eat. Linnaeus gave a brief survey of examples from the animal kingdom, but only provided details concerning the mutualism between the bivalve, Pinna, and the crab, Pinnotheres (Linnaeus 1775:111–113, 1977a). This relationship had been reported by Aristotle (Historia Animalium 547b16–17), but had been neglected by more modern naturalists until Linnaeus’ disciple, Fredrik Hasselqvist (1722–1752), traveled to the eastern Mediterranean (where he died) and confirmed it. Since this Oeconomia naturae dissertation was published in March 1749, and Hasselqvist did not leave Stockholm until 7 August 1749 (Blunt 1971:183–185), Linnaeus obviously added these comments on Pinna and Pinnotheres before the dissertation was republished in Amoenitates Academica, volume 2 (1751), the source of Benjamin Stillingfleet’s English translation. Linnaeus’ survey of the destruction of animals included two food chains, one terrestrial and one aquatic (Linnaeus 1775:114, 1977a):
Next, he emphasized the importance of predators to prevent their prey from over-running everything, and the importance of scavengers to prevent the earth from being overwhelmed with carcasses (Linnaeus 1775:114–122, 1977a). In 1734, while exploring Dalecarlia, Linnaeus had watched his expedition’s horses grazing certain plants and avoiding others. Both John Ray and René Réaumur had reported insects having very specific food plants (Egerton 2005:303 and 2006: ), but in the late 1740s Linnaeus and some students (eight named, plus others) ran 2314 experiments on livestock to determine their plant preferences. Their findings were reported in a dissertation entitled Pan Svecius (Latin 1749, cited from Stillingfleet translation: Linnaeus 1775:361, 1977a):
This was one of the earliest, if not the earliest, series of experiments on an ecological question, and surely the earliest such large-scale quantitative experiments. (Stillingfleet’s translation did not include all details in the original; more is translated in Ramsbottom 1959:162–167.) The reason for so many experiments was that, unlike the insects observed by Ray and Réaumur, these mammals were not limited to eating one or two species, but nevertheless were somewhat selective. Allegedly, God’s reason to make various animal species eat different plant species was to prevent some plant species from becoming extinct due to overeating and others from becoming too abundant because they were not eaten (Linnaeus 1775:347–349, 1977a). Also, in “Oeconomy of Nature”(1775:99–100, 1977a) Linnaeus mentioned “an oeconomical experiment well known to the Dutch,” of which he perhaps learned while he was in the Netherlands in 1735,
In 1774 a dissertation comparable to Pan Svecius appeared on the subject of plants and animals eaten by chickens, ducks, and geese, Esca Avium Domesticarum (Smit 1979:122). Linnaeus’ second most important dissertation for ecology is Politia Naturae (1760), translated into English as ”On the Police of Nature” (1781:129–166). It is on the struggle and survival of species, including humans. A pessimistic conclusion that he drew about humans seemed also to apply to some extent to plant and animal species. Unfortunately, F. J. Brand, the English translator of this dissertation, omitted it. Fortunately, Alan Blair translated it in Kurt Hagberg’s biography of Linnaeus (1952:183).
The point of the dissertation was to explain why a war of all against all (competition) did not lead to extinction. A major reason was what we call ecological diversity: each species is confined to its own “station” (habitat). Sweden had about 1300 plant species, but only about 50–100 are in any one place (Linnaeus 1781:133, 1977b). Linnaeus argued that although it is received opinion that plants were created for the use of animals, actually, animals were created to regulate plants’ abundance. As proof, he cited numerous insect species that only eat a single plant species, doves eat surplus seeds, and other birds, bats, and anteaters eat insects to prevent them from consuming all of the plants they eat, and so on. Linnaeus had a lifelong fascination with insects. Five of the dissertations translated by Brand were on insects and their interactions with other species of plants and animals (Linnaeus 1781:309–456, 1977b). Smit (1979:125) even claimed that Linnaeus made a major contribution to entomology, as evidenced by his Fundamenta Entomologiae (1767). If so, that dissertation was important because it synthesized briefly the works of others. Linnaeus praised as “immortal” the treatise by Réaumur, whom he had met in Paris, and he once sent Réaumur the eggs of the alpicola butterfly (Papilio Apollo). Réaumur’s thank-you note is published in Linnaeus’ correspondence (Smith 1821, II:477–479). However, Linnaeus also pointed out the necessity of providing official names for the species Réaumur had studied (Linnaeus 1772:13). Despite Linnaeus’ strong interest in both insects and plants, he never fully appreciated the role insects play in pollination, except for fig trees. He first named the nectary of flowers in 1735, and he did move from a belief that bees harm flowers by collecting nectar, to a belief that they help pollinate flowers, but never realized their crucial importance for flowers that are not wind pollinated (Miall 1912:322–324, Usinger 1964:6, Lorch 1978:518, 523, Eriksson 1983:105). He did, however, appreciate the danger of accidentally introducing American insect pests when American plants were brought to Europe. In 1739 he confessed to having brought American trees from England to the Netherlands in August 1736; these harbored aphids, which multiplied in a greenhouse and then escaped into botanical gardens in Amsterdam and Leiden (1781:325, 1977b). Pehr Kalm brought pea seeds when he returned to Sweden in 1751; he discovered that they contained live Dermestes pisorum insects, which he captured. However, Linnaeus warned against the danger of artificial introductions (Linnaeus 1781:386–387, 1977). Linnaeus, physician and sometime professor of medicine, followed Richard Bradley (Egerton 2006:124–125) in arguing that minute organisms, “even smaller than the motes dancing in a beam of light”(Linnaeus quoted in translation from Smit 1979:123) transmit contagious diseases. He developed his argument in two dissertations, Exanthemata viva (1757) and Mundus invisibilis (1767). He also discussed and described parasitic worms. His speculations (1973:118–119) about some free-living worms being a different stage of internal parasitic worms was a reasonable hypothesis, but he supported it with unverified (and incorrect) examples (Foster 1965:32, Grove 1990:4, 40, 106, 386). Linnaeus clearly deserves a prominent place among the founders of ecology, but was this a case like Mendel’s, in which his findings were only appreciated after others had rediscovered his basic ideas? No—many of his writings and the student dissertations were reprinted and translated into other languages, and contemporary and later naturalists, including Gilbert White and Charles Darwin, read them (Stauffer 1960, Limoges 1980). Literature cited Blunt, W. 1971. The compleat naturalist: a life of Linnaeus. William Collins and Sons, London, UK. Bowler, P. J. 1989. Evolution: the history of an idea. University of California Press, Berkeley, California, USA. Broberg, G. 2000. Carl Linnaeus, 1707–1777, Swedish naturalist and physician. Pages 417–418 in A. Hessenbruch, editor. Reader’s guide to the history of science. Fitzroy Dearborn, London, UK. Browne, J. 1981. The secular ark: studies in the history of biogeography. Yale University Press, New Haven, Connecticut, USA. Caddy, F. 1886–1887. Through the fields with Linnaeus. Two volumes. Longmans, Green, London, UK. Clements, F. E. 1916. Plant succession: an analysis of the development of vegetation. Carnegie Institution of Washington, Washington, D.C., USA. Du Rietz, G. E. 1957a. Linnaeus as a phytogeographer. Vegetatio 7:161–168. Du Rietz, G. E. 1957b. Linné som myrforskare. (English summary, pages 60–68.) Uppsala Universitets Årsskrift 5:1–80. Egerton, F. N. 1967. Observations and studies of animal populations before 1860: a survey concluding with Darwin’s Origin of Species. Dissertation. University of Wisconsin-Madison, Madison, Wisconsin, USA. Egerton, F. N. 1973. Changing concepts of the balance of nature. Quarterly Review of Biology 48:322–350. Egerton, F. N. 2005. A history of the ecological sciences, part 18: Francis Willughby and William Derham. ESA Bulletin 86:301–313. Egerton, F. N. 2006a. A history of the ecological sciences, part 20: Richard Bradley, entrepreneurial naturalist. ESA Bulletin 87:117–127. Egerton, F. N. 2006b. A history of the ecological sciences, part 21: Réaumur and his history of insects. ESA Bulletin 87:212–224. Ericksson, G. 1983. Linnaeus the botanist. Pages 63–109 in T. Frängsmyr, editor. Linnaeus: the man and his work. University of California Press, Berkeley, California, USA. Foster, W. D. 1965. A history of parasitology. E. and S. Livingstone, Edinburgh, UK. Frängsmyr, T. 1983. Linnaeus as a geologist. Pages 110–155 in T. Frängsmyr, editor. Linnaeus: the man and his work. University of California Press, Berkeley, California, USA. Fries, R. E. 1950. A short history of botany in Sweden. Almqvist and Wiksells, Uppsala, Sweden. Goerke, H. 1993. Linnaeus. D. Lindlley, translator. Charles Scribner’s Sons, New York, New York, USA. Grove, D. I. 1990. A history of human helminthology. C.A.B International, Wallingford, UK. Hagberg, K. 1952. Carl Linnaeus. Alan Blair, translator. Jonathan Cape, London, UK. Hofsten, N. von. 1916. Zur älteren Geschichte des Diskontinuitätsproblems in der Biogeographie. Zoologische Annalen 7:197–353. Hofsten, N. von. 1957. Linnaeus’s conception of nature. Kungliga Vetenskaps– Societetens Årsbok, 65–105. Hulth, J. M. 1907. Bibliographia Linnaeana: matériaux pour server a une bibliographie Linnéenne. [Chronological order.] C. J. Lundström, Uppsala, Sweden. Jackson, B.D. 1913. Authorship in the “Amoenitates Academicae.” Journal of Botany 51:101–103, 244. Kiger, R. W., C. A. Tancin, and G. D. R. Bridson. 1999. Index to scientific names of organisms cited in Linnaean dissertations: together with a synoptic bibliography of the dissertations and a concordance for selected editions. Hunt Institute for Botanical Documentation, Pittsburgh, Pennsylvania, USA. Koerner, L. 1999. Linnaeus: nature and nation. Harvard University Press, Cambridge, Massachusetts, USA. Landin, B.-O. 1972. Charles de Geer (1720–78). Dictionary of Scientific Biography 5:328–329. Larson, J. L. 1971. Reason and experience: the representation of natural order in the work of Carl von Linné. University of California Press, Berkeley, California, USA. Limoges, C. 1974. Economie de la nature et idéologie juridique chez Linné. Thirteenth International Congress for the History of Science (Moscow 1971) Proceedings, Section 9: 25–30. Limoges, C. Dec.1980. De l’économie de la nature aux ecosystemes: l’histoire de l’écologie esquissée à grands traits. Spectre: 9–14. Lindroth, S. 1973. Carl Linnaeus (or von Linné), 1707–1778. Dictionary of Scientific Biography 8:374–381. Lindroth, S. 1983. The two faces of Linnaeus. Pages 1–62 in T. Frängsmyr, editor. Linnaeus: the man and his work. University of California Press, Berkeley, California, USA. Linnaeus, C. 1735. Systema naturae, sive regna tria naturae systematice proposita per classes, ordines, genera, & species. Theodorum Haak, Leiden, The Netherlands. Linnaeus, C. 1737a. Flora Lapponica exhibens plantas per Lapponiam. Salomonem Schouten, Amsterdam, The Netherlands. Linnaeus, C. 1737b. Hortus cliffortianus: plantas exhibens quas in hortistam vivis quam siccis, Hartecampi in Hollandia, coluit virnobillismus & generosissimus Georgius Clifford. Amsterdam, The Netherlands. Linnaeus, C. 1739. Om renarnas brömskulor i Lapland. Kungliga Svenska Vetenskaps Akademien Handlingar 1:121–132. Linnaeus, C. 1740. Anmarking öfwer de diuren, som sagas kåmma neder utur skyarna i norrige. Kungliga Svenska Vetenskaps-Akademien Handlingar 1:326–331. For later editions and translations, see Soulsby numbers 1112–19. Linnaeus, C. 1746a. Fauna svecica sistens animalia Sveciae regni. Salvii, Stockholm, Sweden. Linnaeus, C. 1746b. Oestrus rangiferinus descriptus. Acta Societatis Scientiarum Upsaliensis, for 1741, 102–115. [Linnaeus, C.] 1775. Miscellaneous tracts relating to natural history, husbandry, and physick. To which is added the calendar of flora. B. Stillingfleet, translator. Edition 3. J. Dodsley, Baker and Leigh, London, UK. [Linnaeus, C.] 1781. Select dissertations from the Amoenitates Academicae. F. J. Brand, translator. G. Robinson and J. Robson, London, UK. Linnaeus, C. 1811. Lachesis Lapponica, or a tour in Lapland. C. Troilius, translator. J. E. Smith, editor. Two volumes. White and Cochrane, London, UK. Linnaeus, C. 1964. Systema naturae 1735: facsimile of the first edition with an introduction and a first English translation of the “Observationes.” M. S. J. Engel- Ledeboer and H. Engel, translators. B. de Graaf, Nieuwkoop, The Netherlands. Linnaeus, C. 1971. A tour in Lapland. [Reprint of Linnaeus 1811.] Arno Press, New York, New York, USA. [Linnaeus, C.] 1972. L’équilibre de la nature. B. Jasmin, translator. C. Limoges, introduction and notes. Vrin, Paris, France. Linnaeus, C. 1973. Linnaeus’s Öland and Gotland Journey, 1741. M. Åsbert and W. T. Stearn, translators. W. T. Stearn, introduction. Academic Press, London, UK. [Linnaeus, C.] 1977a. Miscellaneous tracts relating to natural history, husbandry, and physick. B. Stillingfleet, translator. [Reprint of Linnaeus 1775.] Arno Press, New York, New York, USA. [Linnaeus, C.] 1977b. Select dissertations from the Amoenitates Academicae. F. J. Brand, translator. [Reprint of Linnaeus 1781.] Arno Press, New York, New York, USA. Linnaeus, C. 1979. Travels. David Black, editor. Stephen Lee, illustrations. Charles Scribner’s Sons, New York, New York, USA. Lorch, J. 1978. The discovery of nectar and nectarines and its relation to views on flowers and insects. Isis 69:514–533. Mayr, E. 1982. The growth of biological thought: diversity, evolution, and inheritance. Harvard University Press, Cambridge, Massachusetts, USA. Merriam, D. F. 2004. Carolus Linnaeus: the Swedish naturalist and venerable Traveler. Earth Sciences History 23:88–106. Miall, L. C. 1912. The early naturalists: their lives and work (1530–1789). Macmillan, London, UK. Middleton, W. E. K. 1966. A history of the thermometer and its use in meteorology. Johns Hopkins Press, Baltimore, Maryland, USA. Morton, A. G. 1981. History of botanical science: an account of the development of botany from ancient times to the present day. Academic Press, London, UK. Müller-Wille, S. 2003. Nature as a marketplace: the political economy of Linnaean botany. Pages 154–172 in M. Schabas and N. de Marchi, editors. Oeconomies in the age of Newton. Duke University Press, Durham, North Carolina, USA. Nicolas, J.-P. 1963. Adanson, the man. Volume I. Pages 1–121 in G. H. M. Lawrence, editor. Adanson: the bicentennial of Michel Adanson’s “Familles des plantes.” Hunt Botanical Library, Carnegie Mellon University, Pittsburgh, Pennsylvania, USA. Nordenmark, N. V. E. 1935. Anders Celsius, Linné, och den hundragradiga thermometern. Svenska Linné-Sällskapets Årsskrift 18:124–133. Ramsbottom, J. 1959. Caroli Linnaei pan suecicus. Botanical Society of Edinburgh Transactions 38:151–167. Rausing, L. [Koerner]. 2003. Underwriting the oeconomy: Linnaeus on nature and mind. Pages 282–308 in M. Schabas and N. de Marchi, editors. Oeconomies in the age of Newton. Duke University Press, Durham, North Carolina, USA. Schramm, E. 1984. Linné, Gedner und die biologische Kontrolle. Medinin historisches Journal 19:244–249. Smit, P. 1979. The zoological dissertations of Linnaeus. Svenska Linnésällskapets Årsskrift for 1978, 118–136. Smith, J. E., editor. 1821. A selection of the correspondence of Linnaeus, and other naturalists, from the original manuscripts. Longman, Hurst, Rees, Orme, and Brown, London, UK. Soulsby, B. H. 1933. A catalogue of the works of Linnaeus (and publications more immediately relating thereto) preserved in the libraries of the British Museum. [Topical order.] Edition 2, British Museum, London, UK Spary, E. C. 2002. Carolus Linnaeus (Carl Linnaeus, Carl von Linné:1707–78), Swedish naturalist. Volume 2. Pages 410–413 in A.C. Kors, editor. Encyclopedia of the Enlightenment. Oxford University Press, Oxford, UK. Stafleu, F. A. 1971. Linnaeus and the Linnaeans: the spreading of their ideas in systematic botany. Oosthoek, Utrecht, The Netherlands. Stauffer, R. C. 1960. Ecology in the long manuscript version of Darwin’s Origin of species and Linnaeus’ Oeconomy of nature. American Philosophical Society Proceedings 104:235–241. Usinger, R.L. 1964. The role of Linnaeus in the advancement of entomology. Annual Review of Entomology 9:1–16. Acknowledgments For their assistance I thank Jean-Marc Drouin, Muséum National d’Histoire Naturelle, Paris; and Anne Marie Drouin-Hans, Université de Bourgogne. Frank N. Egerton
|
||||||||||||||||||||||||||||||||
|
Serving Society Through Ecological Expertise Autumn 2006 |
|
* At ESA's Annual Meeting in Memphis, TN...
Media Activities... ESA's Public Affairs Committee and RRT Congressional Staff ... ESA often hosts one or two congressional staff members at the Annual Meeting, enhancing the communication of science and establishing personal networks between scientists and policy-makers. The congressional staff member hosted in Memphis met with Agroecology RRT members as well as RRT member and Vice-President for Public Affairs Rich Pouyat. RRT Luncheon ... ESA's second annual luncheon for RRT members highlighted the accomplishments of the RRTs over the past two years and featured a working discussion led by outgoing ESA President nancy Grimm on the future RRT structure and functions.
|
|
| * Looking Ahead... ESA's Public Affairs Office is planning the next round of RRT activities, and working on enhancing and updating ESA's Expert Guide and web site. Stay tuned—we will be contacting you for your feedback and updated biographical information. Please send ideas and suggestions for future RRT opportunities and activities to Laura Lipps at [email protected]. | |
* Providing Input to the Media ... |
|
* Supporting Science at Federal Agencies ... Providing Comments on Rulemaking... ESA's President, with the assistance of Aquatic Ecology RRT members, submitted comments to the Environmental Protection Agency on its proposed rule on stream mitigation. The comments are available at: Supporting Funding for Science ... RRT members Alan Townsend joined 80 other scientists to discuss science funding with Members of Congress, as an ESA representative for the Coalition for National Science Funding, ESA's Public Affairs Office brings RRT members, students, and scientists from all career stages to D.C. for science policy training and targeted meetings with Congressional offices. |
Public Affairs Perspective
|
ESA Statement on Massachusetts et al. vs. U.S. Environmental Protection Agency,
|
Note: Dr. Harold Ornes is the editor of Ecology 101. Anyone wishing to contribute articles or reviews to this section should contact him at the Office of the Dean, College of Science, Southern Utah University, 351 W. Center, Cedar City, UT 84720; (435) 586-7921; Fax: (435) 865-8550; E-mail: [email protected]. Whether you are teaching undergraduate or graduate-level field ecology, or you are a full-time researcher and in the midst of gleaning the maximum amount of information from your most recent field research sampling trip, I think you will agree that we almost never remember everything needed in the field, and likewise almost never have a perfectly smooth and uneventful field trip or season. Thanks to Carmen Wong, University of British Columbia, we can take a few moments, sit around the fire, feel her pain, and start planning for next season. I would encourage you to share one or two of your field season horror stories with your peers through this column. |
Preparing for the Field SeasonI thought I was entering the field season for my Ph.D. research in good shape. I had over 5 years of experience as a consultant working on various field research projects. And, I had spent time in the field during my Master’s research ziplocking grizzly bear feces in the Yukon. So, as a seasoned field biologist embarking on my Ph.D. research, I finally had my research permit in place, lined up two great field assistants, and purchased several boxes of waterproof paper and printed colored maps of my potential study sites. But, I should have known that you can never be too prepared for fieldwork. My first challenge was losing a field assistant on the first day. I didn’t actually lose him in the forest; rather, he took a permanent job with a consulting company. My promise of helicopter rides in the Rockies could not compare to a decent salary with health benefits. I spent the next month looking for another field assistant. It turns out the good candidates get snapped up early. My second challenge was with my sampling design. I had wanted to use stratified random selection of stands containing whitebark pine in order to describe dynamics across large landscapes. Random selection using a vegetation inventory and GIS had worked in a pilot study the year before in another park. Using my beautiful colored maps, my remaining field assistant and I spent a week bushwhacking to mountaintops, only to find no whitebark pine where it was suppose be. It seems that I was mistaken in my assumption that I was using a vegetation inventory. An inventory did not exist and I was actually using a coarse-grain classification. Polygons indicated only a potential for containing whitebark pine … and clearly that potential was low. It turns out that having the home number of a biometrician is handy when one is revising a sampling design in the field. My third challenge was in vexing the park wardens. We left the door of a warden cabin in the backcountry unlocked. We almost lost the privilege of using these cabins, which would have made my research impossible, but luckily the supervisory warden was understanding. It was clear, however, that a moment of carelessness can jeopardize one’s research. So what should you walk away with after reading the above tales? Here are a few lessons I learned this summer that may help you during your next field project:
Finally, get out there and enjoy yourself. If you aren’t enjoying what you are doing, why are you doing it? Carmen Wong |
Since the Bulletin has gone electronic, the hope is that our journal will fill the niche between the science papers of Ecology and Ecological Applications and the science/policy of Frontiers in Ecology and the Environment. One area that has been suggested by readers is essays on papers that particularly influenced the ideas and approaches to ecology of ecologists who have contributed to that particular subdiscipline of ecology. The idea is that the essay will both identify a paper or papers that were particularly important in catalyzing future developments, at least to the writer of the essay, and will also be a personal viewpoint. The working title for this section is: “The Paper Trail.” The first in this new series is Steward T. A. Pickett’s essay on W. S. Cooper’s classic paper on plant succession. The original paper can be viewed at: “ The fundamentals of vegetational change, ” William S. Cooper, Ecology 7(4):391–413, October 1926), Stable URL: ‹http://links.jstor.org/› |
The Paper Trail: W. S. Cooper’s “Fundamentals of Vegetational Change” and a Fluent Mode of Thought for EcologyIt is a bit amazing to admit that a paper that influenced me is now 80 years old! I encountered Cooper (1926) first in graduate school, but I have revisited it at intervals, and have been repeatedly stimulated and refreshed by it. In spite of the fact that I originally consulted this paper for its insights into plant succession, it is important not so much for what is says about succession, but for the analytical approach it takes. I believe the approach is one that can benefit ecologists in any specialty and at any age. First a little background is in order. William Skinner Cooper, a professor of Botany at the University of Minnesota, was one of the most important of the first generation of American ecologists (McIntosh 1985). If, by chance, you are an American plant ecologist, you may well trace your lineage back to him (Sprugel 1980). He was the major professor of a number of active members of the second generation, who in turn trained many academic grand- and great-grandchildren. Henry J. Oosting, Murray F. Buell, and Rexford Daubenmire were especially prolific members of the second generation. This legacy is certainly a noteworthy role in the field. But when one looks at the big debate of the day—the nature of community dynamics—Cooper is conspicuous in not holding a dogmatic view about succession, or in unfailingly supporting either Clements or Gleason, whose differences have come to symbolize the debate. Instead, Cooper was guided by careful observation of communities in the field, and was remarkably free of ideology. The paper under consideration perhaps explains his open-mindedness and ability to see beyond ideology and ossified positions. Here is what “The fundamentals of vegetational change” (Cooper 1926) did for me. 1 Cooper opens with a goal of examining and constructively criticizing the basics of his discipline: “A periodic inspection of foundations is most desirable for any edifice, and particularly so when the superstructure is being continually added to, as in the development of scientific knowledge.” (Cooper 1926:391). This is challenging stuff, because at that time it had been a mere 10 years since the publication of Clements’ (1916) big book on succession, with its proliferation of terminology, and exclusion of special cases, that promoted the questionable “organismal” concept of the plant community. Of course, the assumption that communities or ecosystems are equivalent to organisms in their development and stabilization has long since been rejected (Botkin and Sobel 1975). At the time, however, it was a persuasive and widely held idea. Cooper is telling us, whatever the orthodoxy, to go back to basics, and see what should stay and what should be rejected. This is the deepest lesson of his paper, which invites each generation of ecologists to reexamine the foundations of their discipline. What should a rigid and dogmatic classification of succession, or for that matter, any ecological phenomenon, be replaced with? Oddly enough, although the study of succession and community change was called “dynamic ecology” at the time, Cooper noted that there was a proliferation of static classification systems of succession, each type having a narrow set of specific causes associated with it. He was concerned that the rigidity of classification systems would force ecologists to put actual communities in boxes that were not appropriate, and at any rate, that the classifications did not really account for change. He said, “It is our task to give verbal expression to constantly changing phenomena in a way that will parallel their mutations as closely as humanly possible.” (Cooper 1926:396). This was a call for flexible concepts and terms that reflected the flexibility—or perhaps in today’s terms, complexity—of the world to which the concepts referred. Such flexible terms would emerge from the actual process itself, rather than emerging from an idealization of it. This meant that “We must, accordingly, rigorously exclude all stock ecological terms and phrases, and treat the phenomena in a purely descriptive manner.” (Cooper 1926:396). So clear articulation of what the core phenomenon is, separate from the assumptions about it that yield idealized patterns or processes, becomes a key intellectual activity in ecology. The specific features that Cooper identified as fundamental in succession are less important for the story I am telling than the approach he used. However, his summary of the fundamentals proved helpful to me in sorting through the problems of succession that were becoming clear in the 1970s, when I first encountered his paper (Box 1). For example, he took as fundamental that change in vegetation is universal, and included both contemporary and paleoecological changes. Change in vegetation could be due to any cause, not just ones that were expected to make succession move “forward.” Cooper, furthermore, included causes that arose from the characteristics and interactions of plants themselves, as well as those that were external to plants. Such things as soil differences in a biome, or fire, or people were all acceptable causes to Cooper, although Clements and his adherents viewed them as exceptions to be excluded from the study of succession. He took the interaction of plants and environmental causes (including animal activities) as a system. His recognition that different causes acted on various scales seems quite prescient of contemporary interest in scale. Cooper also indicated that all causes are always acting on vegetation change, though to different degrees. It was the complex interaction of a broad array of causes that resulted in vegetation change.
This insight of Cooper’s concerning the broad applicability of causes, appears later in the idea that net effects in succession reflect an array of mechanisms. A hierarchical set of causes, which may act differentially at different periods of succession, is one way to apply Cooper’s insight (Pickett and Cadenasso 2005). Also important to me was the insight that “Destruction is inherent in all successions” (Cooper 1926: 404), which of course helped me recognize the broad role of natural disturbance in communities (Pickett and White 1985). This was not the only paper of Cooper’s to make that point. Cooper’s concern with all causes and scales of vegetation dynamics was an important exemplar for me. He sought principles to explain all sorts of vegetation change, ranging from evolution on one extreme, through the grand shifts of paleoecology, to detailed contemporary community alteration. Therefore, his work suggested a broad and inclusive framework that could account for the evolutionary basis of succession (Pickett 1976), to the role of animals, herbivores, and predators (which had long been neglected by plant ecologists). The pursuit of broad, open-ended, and intentionally mutable frameworks has become one of the tasks I have set for myself, in part due to the insights offered by Cooper so long ago. Cooper’s fluid thinking was easy for me to apply to vegetation dynamics, since that field initially stimulated his essay. However, his charge for each generation to examine the fundamentals of its concepts and frameworks echoes throughout ecology. Rigid classificatory systems of ecological phenomena, inappropriate reification of idealized concepts, and narrow conceptions of trajectories and causes, is not uncommon in ecology. Cooper’s exhortation for us all, whatever speciality we practice, to periodically examine our fundamentals, is as cogent today as when he put pen to paper over 80 years ago.
I hope that the emerging and tentative frameworks for vegetation change (Pickett and Cadenasso 2005), boundary dynamics (Cadenasso et al. 2003), landscape heterogeneity, and patch dynamics (Pickett et al. 2000), show the mark of Cooper’s influence, and at their best, rise to the level of fluid thinking for fluid phenomena. In any event, his call to reevaluate the fundamentals seems as relevant to all parts of ecology today as his original call that emerged from vegetation dynamics in 1926. Botkin, D. B., and M. J. Sobel. 1975. Stability in time-varying ecosystems. American Naturalist 109:625–646. Cadenasso M. L., S. T. A. Pickett, K. C. Weathers, and C. G. Jones. 2003. A framework for a theory of ecological boundaries. BioScience 53:750–758. Clements, F. E. 1916. Plant succession: an analysis of the development of vegetation. Carnegie Institution of Washington, Washington, D.C., USA. Cooper, W. S. 1926. The fundamentals of vegetational change. Ecology 7:391–413. McIntosh, R. P. 1985. The background of ecology: concept and theory. Cambridge University Press, Cambridge, UK. Pickett, S. T. A. 1976. Succession: an evolutionary interpretation. American Naturalist 110:107–119. Pickett, S. T. A., and M. L. Cadenasso. 2005. Vegetation succession. Pages 172–198 in E. van der Maarel, editor. Vegetation ecology. Blackwell Publishing, Malden, Massachusetts, USA. Pickett, S. T. A., M. L. Cadenasso, and C. G. Jones. 2000. Generation of heterogeneity by organisms: creation, maintenance, and transformation. Pages 33–52 in M. Hutchings, editor. Ecological consequences of habitat heterogeneity. Blackwell, New York, New York, USA. Pickett, S. T. A., and P. S. White, editor. 1985. The ecology of natural disturbance and patch dynamics. Academic Press, Orlando, Florida, USA. Sprugel, D. G. 1980. A “pedagogical genealogy” of American plant ecologists. ESA Bulletin 61:197–200. Steward T. A. Pickett 1 Rereading Cooper’s paper in 2006 raises two questions about style. His use of “man” rather than “people” or “humans” is something that I find jarring. While I realize that grammarians may argue that is an acceptable use, I must warn contemporary readers that they may have to read beyond that usage. Cooper’s style is otherwise just a little more formal than today’s most formal writing. It is remarkably clear writing in spite of that formality. |
|
Upstart Views of Restoration Icons
As ecological restoration efforts become better known and better studied, and as more ecologists choose to conduct research in restoration sites, the gap between applied science and theory becomes narrower. Such was the case when Margaret Palmer catalyzed a symposium for the 2002 ESA meeting in Tucson, where ecologists indicated how theory could inform restoration practice. Several speakers then expanded their remarks in a book (Falk et al. 2006) that further merged ecological theory and practice. Still, gaps will remain if the icons relied upon by restorationists are inconsistent with the latest advances in theory and understanding of how ecosystems develop. Accordingly, this symposium set out to deconstruct some icons of restoration ecology, by applying contemporary ecological theory to restoration ecology. The combined result represents a paradigm shift, away from equilibrial, predictable models and toward a world of restored ecosystems that is at least partly stochastic, time-varying, and context dependent. One pervasive icon is a simple illustration of how ecosystem structure and function develop following some restoration effort (Fig. 1). A. D. Bradshaw (1984) depicted restoration as having a single target that is hit after substrates are modified, with ecosystem structure and function developing along the same linear pathway. His diagram continues to appear in books, papers, and even the journal Science (Dobson et al. 1997), although evidence supporting this model has never been included in those publications (Zedler and Lindig-Cisneros 2000). Our concern is not with Tony Bradshaw, who is himself an important icon of restoration ecology, for he was among the earliest ecologists to address actual restoration problems (how to vegetate mine tailings) and to urge others to recognize the value of studying restoration sites as a test of ecological understanding (e.g., Bradshaw 1987). Instead, the issue concerns those who expect restoration efforts to follow overly simple models, or who promise unrealistic pathways and outcomes.
Because there is no complete guide to the theoretical foundations of restoration ecology, and because ecologists have much to offer in helping restoration ecology mature as a science (Zedler 2005), we convened a symposium on “Upstart Views of Restoration Icons” to highlight some of contributions of the new book (Falk et al. 2006), while asking authors and other speakers to stretch beyond previous writings to close further the gap between theory and practice. Dan Larkin solicited the speakers and organized the session; Joy Zedler moderated the symposium; and Don Falk provided the wrap-up. As a disclaimer, we did not ask speakers to critique the icon in Fig. 1; rather, we selected results from their talks that address this widely published model of how restoration works; thus, our descriptions of each presentation are not fully representative (in keeping with instructions for symposium commentaries in the ESA Bulletin). Ten speakers (names in boldface type) provided evidence that dispels key elements of a key restoration icon (Fig. 1). Their presentations failed to support (1) a single obvious target for restoration, (2) a straight path to the target, and (3) a linear relationship between structure and function. Upstart view No. 1. There is no single, obvious target for restoration; that is, reference conditions represent dynamic, multivariate, and nonequilibrial processes. Bob Peet (University of North Carolina) presented a model of relationships between environmental conditions and plant communities that suggests appropriate species lists, proportions of plants/species, and sequences for introducing plants to individual restoration sites. Science-based “designer plantings” should reduce restoration efforts and costs. North Carolina’s Department of Transportation is using this approach to restore lands in anticipation of the need to mitigate impacts of future highway construction projects. Roberto Lindig-Cisneros (University of Michoacana) added human needs to the list of constraints on the restoration target. In southern Mexico, managers agreed to allow experimental restoration of tephra (unvegetated ash that persisted 60 years following eruption of Mt. Paracutín), but only if the target could be the two native pine species that provide livelihoods (Fig. 2). In exchange, local people helped establish experimental plantings (with and without bark mulch). As expected, mulching lowered soil temperature and enhanced pine establishment, but, unexpectedly, the effect was strong only in dry years.
Denise Seliskar (University of Delaware-Lewis) demonstrated that tailoring could extend below the species level. After planting a salt marsh restoration site in Delaware (Fig. 3, photo) as an experiment to test the effects of three genotypes of Spartina alterniflora (from Maine, Delaware, and Georgia), she and others documented numerous impacts on everything from canopy height and stem density (Fig. 3, graphs), to root and rhizome distributions, edaphic chlorophyll concentration, and decomposition rates. Even the numbers of larval fish caught in pit traps differed by a factor of 2. She and Jack Gallagher then extended their research by selecting genotypes of many halophytes via tissue culture. Some genotypes are broadly tolerant of stressful field conditions, while others perform best in specific sites. “Designer genotypes” could increase survival and growth.
Stuart Findlay (Institute for Ecosystem Studies, New York) gave compelling evidence that multiple-function ecosystems are unrealistic targets for wetlands. While we might aim for clean water, high productivity, high biodiversity, flood reduction, and other functions, he argued that restoration sites are context dependent and typically do not provide all the ecosystem services expected of them, and that some combinations of functions are mutually exclusive (Fig. 4). If multiple driving factors are not correlated in space or time, individual sites cannot excel in several functions simultaneously. This leads to high interannual variability in site performance and argues for a relaxation of the restoration target. Thus, plans to restore multiple functions in a watershed will require multiple restoration sites and efforts.
As Don Falk concluded, the paradigm of the “single target” should shift to that of “reference dynamics” (Falk, in press), where interactions, temporal and spatial variability, and stochastic processes are emphasized. This makes defining the ecological reference more complex, difficult, and uncertain—but also more realistic.
Katie Suding (University of California-Irvine) offered evidence that California grasslands resist restoration and follow an alternative states model (Fig. 5), with internal feedbacks that help sustain each of the dominance states, namely, the native bunch grass (which competes more strongly for N) and exotic annual grasses (which compete more strongly for light). Through innovative experimentation, she showed that soils formed under the native tended to favor the native, and that soils formed under exotics favored exotics. These and other results collectively support the alternative state model. Thus, if restorationists plant the “right” species in the “wrong” soil, the site will not necessarily favor the native vegetation.
Dan Larkin (University of Wisconsin) tested the importance of intertidal pools and tidal creeks to salt marsh food webs in a large experimental salt marsh (Fig. 6) and found increased feeding opportunities at high tide (more algal biomass, more invertebrates in pools) than where pools were lacking. Also, killifish fed more in areas with pools than without. Furthermore, tidal creeks enhanced use by mudsuckers. Thus, feeding opportunities were best restored where topography mimicked natural heterogeneity (creeks plus pools), but the relationships between species and microhabitat were complex.
Holly Menninger (University of Maryland) culled the literature for examples of real evidence that substrate heterogeneity increases the diversity of taxa. In contrast to the above speakers, she found none for a dozen experiments in streams, including the work of her co-author, Margaret Palmer (Fig. 7). While the experiments being done in streams have not enhanced diversity of the taxa being explored, a focus on restoring processes in streams might help researchers figure out how to enrich diversity.
As Don Falk summed it up, the emerging view of the postrestoration state is one of complex, and at least partly stochastic, spatially contingent systems, with nonlinear response functions to treatments, nondeterministic outcomes, and nonequilibrial properties. Upstart view No. 3. Structure and function are not linearly related. Shahid Naeem (Columbia University) indicated that many plant species combinations might be viable restoration targets, based on biodiversity–ecosystem function (BEF) theory, and that the relationship to species richness will differ by function. He unveiled a new 5-year experiment (Fig. 8) that is underway in Mongolia, involving >700 plots (each 6 × 6 m) that will have controlled composition and species richness for the first years, before opening plots to grazing and assessing their functional capacity (including livestock production). “Combinatorial forecasting” could lead to recommendations for specific richness levels and/or assemblages for use in restoring specific ecosystem functions.
Greg Bruland (University of Hawaii) tested the ability to restore both species and functions to a former forested wetland that was farmed (and flattened in the process), and then regraded to create a large experiment in topographic heterogeneity (1.3-m vertical range). Aboveground biomass accumulated least on hummocks and most in hollows, while species richness increased from hummocks to hollows to flats (Fig. 9)—not a linear correlation between these measures of function and structure. Overall, the specialization of species to microhabitats led to high diversity at the site scale.
Joy Zedler (University of Wisconsin) filled a gap in the program with Suzanne Kercher and Andrea Herr-Turoff’s data from wet prairie mesocosms. As an invasive grass expanded, biomass increased and species richness decreased. Then, despite killing the grass with herbicide, biomass remained high where few species remained. Productivity and species richness were negatively related, contradicting the notion of a positive linear correlation; furthermore, restorability was lower where stormwater treatments (flooding and nutrients) were continued than where they ceased. Clever theorists and talented experimentalists continue to amass impressive data sets that challenge traditional views that restoration outcomes are predictable (we achieve a specific target), defined (variation is around a predetermined mean condition), and stable (we can keep the system that way). We summarize with: Upstart view No. 4: Restoration outcomes are unpredictable, stochastic, and nonequilibrial, and the work is never finished. While the “Bradshaw icon” might characterize some components of some restoration projects at some times, a more dynamic paradigm for restored ecosystems would accommodate variability and even unpredictability as positive signs of healthy, functioning systems. Literature cited Bradshaw, A. D. 1984. Ecological principles and land reclamation practice. Landscape Planning 11:35–48. Bradshaw, A. D. 1987. Restoration: an acid test for ecology. Pages 23–29 in W. R. Jordan III, M. E. Gilpin, and J. D. Aber. Restoration ecology: a synthetic approach to ecological research. Cambridge University Press, Cambridge, UK. Brooks, S. S., M. A. Palmer, B. J. Cardinale, C. M. Swan, and S. Ribblett. 2002. Assessing stream ecosystem rehabilitation: limitations of community structure data. Restoration Ecology 10: 156–168. Bruland, G. L., and C. J. Richardson. 2005. Hydrologic, edaphic, and vegetative responses to microtopographic reestablishment in a restored wetland. Restoration Ecology 13:515–523. Dobson, A. P., A. D. Bradshaw and A. J. M. Baker. 1997. Hopes for the future: restoration ecology and conservation biology. Science 227:515–522. Falk, D. A. In press. Process-centered restoration and reference dynamics. Journal of Nature Conservation. Falk, D., M. Palmer, and J. B. Zedler, editors. Foundations of restoration ecology. Island Press, Washington, D.C., USA. Seliskar, D. M, J. L. Gallagher, D. M. Burdick, and L. A. Mutz. 2002. The regulation of ecosystem functions by ecotypic variation in the dominant plant: a Spartina alterniflora salt-marsh case study. Journal of Ecology 90:1–11. Zedler, J. B. 2005. Restoration ecology: Principles from field tests of theory. San Francisco Estuary and Watershed Science. ‹ http://repositories.cdlib.org/jmie/sfews/vol3/iss2/art4› Zedler, J. B., and R. Lindig-Cisneros. 2001. Functional equivalency of restored and natural salt marshes. Pages 565–582 in M. Weinstein and D. Kreeger, editors. Concepts and controversies in tidal marsh ecology. Kluwer Academic, Dordrecht, The Netherlands. Joy B. Zedler, Botany Department and Arboretum, University of Wisconsin-Madison, Madison, WI 53706, E-mail: [email protected] Donald A. Falk, Laboratory of Tree-Ring Research, University of Arizona, Tucson, AZ 85721 Daniel J. Larkin, Botany Department, University of Wisconsin-Madison, Madison, WI 53706 |
Urban Food Webs: Errata. Revised Text and Figures Urban Food Webs: Errata. Revised Text and Figures In the October 2006 issue of the ESA Bulletin, two errors appeared in the Symposium Review “Urban Food Webs” (pages 387–393). In the list of authors, Thomas Parker should have appeared as Tommy Parker. And in Fig. 3a (page 389), the scientific name should be spelled “Habrophlebiodes,” as shown in the corrected figure below.
|
|
Canada Chapter NewsletterChapter officers Announcing the new ESA Canada Chapter web site! Please visit our newly revamped web site which you can find at: ‹http://www.esa.org/canada/›. Here you will find items of interest to Canadian ecologists including announcements of upcoming meetings. We have also set up a blog to discuss current Canadian environmental policy issues. We are currently working on the new look for the web site. If you are interested in making suggestions for content or design, please contact Karen Yee at: [email protected] or (403) 220-5264. Any West Coaster interested in representing the Canada Chapter?? We are looking for someone who is willing to represent the ESA Canada Chapter at the Pacific Ecology and Evolution Conference in March 2007, which has been sponsored by our chapter. If someone is planning on attending the Conference and would like to help the chapter out, please contact Karen Yee at: [email protected] or 403-220-5264. Minimal work is required; we just want some representation at the meeting.
We are pleased to announce that the second Canadian Society of Ecology and Evolution (CSEE) Annual Meeting will be held in Toronto, 17–20 May 2007 at Victoria College, University of Toronto. There will be three plenary talks, three symposia, 132 contributed talks, two poster sessions, and rooms for evening workshops. We expect about 500 delegates. More information can be found at: ‹http://www.eeb.utoronto.ca/csee/index.html›. Respectfully submitted, Southeastern Chapter Newsletter
|

|
 |
The Ecological Society of America's For more information visit ‹http://www.esa.org/sanjose/› |
|
The conference will be held 18–20 June 2007 at the World Forestry Center, Portland, Oregon, USA. As our use of land for nonnative and nonagricultural purposes increases, traditional wildlife habitats are lost. As a result, wildlife managers, ecologists, city planners, landscape architects, and community leaders must find ways to create and maintain ecologically significant habitats in and around urban areas. This conference demonstrates management opportunities for urban habitats from small city parks in urban centers to large subdivisions with considerable open space. The roles of urban soils, vegetation, water quality, noise, open space, greenways, habitat, animal communities, and human communities will be discussed. Presenters will share proven strategies for successfully integrating increased wildlife habitat in our cities and towns. For further information contact: The National Arbor Day Foundation
|
|
Instructions for ContributorsDEADLINES: Contributions for publication in the Bulletin must reach the Editor’s office by the deadlines shown below to be published in a particular issue: |
Issue
January (No. 1)
April (No. 2)
July (No. 3)
October (No. 4)Deadline
15 November
15 February
15 May
15 August
Please note that all material for publication in the Bulletin must be sent to the Bulletin Editor. Materials sent to any address except that of the Editor, given below, must then be forwarded to the Editor, resulting in delay in action on the manuscripts. Send all contributions, except those for Emerging Technologies, Ecology 101, Ecological education K–12, and Obituaries/Resolutions of Respect (see addresses below), to E .A. Johnson, Bulletin Editor-in-Chief, Department of Biological Sciences, University of Calgary, Calgary, Albert, T2N 1N4 Canada. Phone (403) 220-7635, Fax (403) 289-9311, E-mail: [email protected].MANUSCRIPT PREPARATION: The manuscript should be submitted as a WordPerfect or Microsoft Word (for Mac or DOS) manuscript, preferably as an e-mail message attachment to [email protected]. E-mailed photographs and diagrams must be in .tiff or .eps format. Other forms of electronic copy (text embedded in e-mail messages, diskettes sent by post) or hard copy can be submitted if absolutely necessary. If formatting could be troublesome (e.g., tables, European alphabet characters, etc.), hard copy also should be sent via fax to E. A. Johnson at (403) 289-9311, or via post. Hard-copy manuscripts should be double-spaced, with ample margins. Plain formatting must be used on hard-copy and electronic manuscripts. PLAIN FORMATTING consists of a single font of a single size, left justification throughout, line spacing the same throughout, and up to three different weights of headings. Other formats will not be accepted for publication. The author should THOROUGHLY PROOF the manuscript for accuracy, paying special attention to phone and fax numbers and web site and e-mail addresses, which are frequently incorrect.COVER PHOTOGRAPHS: The photo should illustrate ecological processes or an ecological research design. The cover of the July, 2004 issue is a good example. It helps if the colors in the photo are bright, although black and white photos are considered if they are well composed with good contrast.
If you would like to submit a digital file, submissions can be small jpegs (72 dpi) but if the image is selected for a cover the final image must be 300 dpi and at least 7 inches wide and 5 inches high. Email the file as an attachment to the Editor of the ESA Bulletin at [email protected]. Or send a single 5 x 7 or 8 x 10 photo to the Bulletin. On an accompanying photocopy, give your name, address, a photo legend up to 100 words, and, if the photo describes a paper in ESA or in another journal, the literature citation or title of the accepted manuscript. If you wish unused photos to be returned please include a self-addressed return envelope.LETTERS TO THE EDITOR AND COMMENTARIES: Please indicate if letters are intended for publication as this is not always obvious. The Bulletin publishes letters, longer commentaries, and philosophical and methodological items related to the science of Ecology. There are no page limits but authors may be asked to edit their submissions for clarity and precision. Previously published items from other sources can be republished in the Bulletin if the contributor obtains permission of the author and the copyright holder, and clearly identifies the original publication.MEETING ANNOUNCEMENTS: Submit a brief prose description of the upcoming meeting, including title, a short paragraph on objectives and content, dates, location, registration requirements, and meeting contact person’s name, street address, and phone/fax/e-mail address. Please do not submit meeting brochures in the expectation that the Editor will write the prose description; he won’t. Compare the publication deadlines above with the meeting deadlines to be sure the announcement will appear in time.MEETING REVIEWS: The Bulletin publishes reviews of symposia and workshops at the annual ESA meeting, as well as important and appropriate meetings that are unrelated to the annual ESA meeting. The reviewer should strive for a synthetic view of the meeting or symposium outcome, i.e., how the various presentations fit or conflict with each other and with current scientific thought on the topic. Review length is open, although about four double-spaced pages should be enough to capture the essence of most meetings.
The following advisory items are provided to help focus your review.
a) Meeting title, organizer, location, sponsoring organizations?
b) What were the meeting objectives, i.e., what scientific problems was the meeting organized to solve? Who cares (i.e., what was the relevance of this scientific problem to related ones under examination)?
c) How well did the meeting meet the objectives? Were there specific papers delivered or roundtables/discussion groups that were exemplary in reaching the objectives? You may concentrate the review on only the outstanding papers to the exclusion of all others, or give a comprehensive view of all presentations/meeting activities, or examine a selection of papers that neither describes all, nor focuses on a very few.
d) What new was discussed? What previously weak hypotheses were strengthened, confirmed or supported? Were any breakthroughs, or new or innovative hypotheses presented, that forced participants to rethink current concepts?
e) Was there anything else important that the meeting accomplished that may not have been part of its explicit objectives?
f) What subjects relevant to the meeting objectives were missing or left out? Did the scientific components of the problem that were included produce a strong slant or serious void by virtue of blind spots by the organizers, failure of invitees to appear, or similar difficulties?
g) Are there plans for a proceedings issue or meeting summary document, and if so who is editing it, who is publishing it, and when is it planned to appear (i.e., where can interested folks learn more about the meeting?)EMERGING TECHNOLOGIES: Submissions for this section should be sent to the Section Editors in charge of the section: Dr. David Inouye, Department of Biology, University of Maryland, College Park, MD 20742. E-mail: [email protected]; or Dr. Sam Scheiner, Div. of Environmental Biology, Natl. Science Foundation, 4201 Wilson Blvd., Arlington, VA 22230. E-mail: [email protected]ECOLOGY 101: Submissions should be sent to the Section Editor in charge of this section: Dr. Harold Ornes, College of Sciences, SB 310A, Southern Utah University, Cedar City, UT 84720. E-mail: [email protected]ECOLOGICAL EDUCATION K–12: Correspondence and discussions about submissions to this section should be sent to Susan Barker, Department of Secondary Education, 350 Education South,, University of Alberta, Edmonton, Alberta T6G 2G5 Canada. E-mail: [email protected]
(780) 492 5415 Fax: (780) 492 9402
or
Charles W. (Andy) Anderson, 319A Erickson Hall, Michigan State University, East Lansing, MI 48824 USA. E-mail: [email protected]
(517) 432-4648 Fax: (517) 432-5092FOCUS ON FIELD STATIONS: Correspondence and discussions about submissions to this section should be sent to E. A. Johnson, Bulletin Editor-in-Chief, Department of Biological Sciences, University of Calgary, Calgary, Albert, T2N 1N4 Canada. Phone (403) 220-7635, Fax (403) 289-9311, E-mail: [email protected].OBITUARIES AND RESOLUTIONS OF RESPECT: Details of ESA policy are published in the Bulletin, Volume 72(2):157–158, June 1991, and are abstracted below. The death of any deceased member will be acknowledged by the Bulletin in an Obituary upon submission of the information by a colleague to the Historical Records Committee. The Obituary should include a few sentences describing the person’s history (date and place of birth, professional address and title) and professional accomplishments. Longer Resolutions of Respect, up to three printed pages, will be solicited for all former ESA officers and winners of major awards, or for other ecologists on approval by the President. Solicited Resolutions of Respect will take precedence over unsolicited contributions, and either must be submitted to the Historical Records Committee (see ESA website) before publication in the Bulletin.