Ecological Archives E096-260-A1
Nathan J. Dorn and Mark I. Cook. 2015. Hydrological disturbance diminishes predator control in wetlands. Ecology 96:29842993. http://dx.doi.org/10.1890/14-1505.1
Appendix A. Additional methodological detail.
A. Additional description of the Loxahatchee Impoundment Landscape Assessment (LILA) wetlands and sampling methods.
The LILA wetlands are situated immediately east of and adjacent to the Arthur R. Marshall Loxahatchee National Wildlife Refuge (Water Conservation Area 1) in Palm Beach County Florida. Each wetland macrocosm has a deep distribution pond on the west side (Fig. A1).; Water is added to the distribution ponds through a gated culvert from a header cell. A few smaller deep water (> 2 m in summer) habitats are present on the perimeters. The water level in each wetland can be independently set by management of the distribution header cell, the inflow gate (west side) and outflow gate (east side). The entire system re-circulates through a series of canals and has no surface flow connections with other canals or wetlands. M1 and M3 are managed as non-flowing wetlands similar to the condition of impounded wetlands in southern Florida. M2 and M4 are managed as slowly flowing (~ 1 cm sec-1) wetlands, especially throughout the wet season (more typical of the historical Everglades). The flowing conditions of M2 and M4 mean that fish populations are more connected to each other (through canals/header cell) than M1 and M3 (i.e., gates are mostly shut except when water levels are being changed for a day or night). Each macrocosm has a larger deep slough and a smaller shallow slough running the length of the wetland (Fig. A1, Aich et al. 2011) and our sampling work was focused on those habitats at times when the shallower vegetated ridges were largely dry (0–10 cm water depth). Four small alligator ponds (~7 m diam. each) were present in each wetland to be representative of natural deep water habitats. Other deep water ditches on the ends and sides of the macrocosms represent an unnatural habitat for the Everglades and while deep water (alligator holes) make up ≤ 0.015% of the available habitats in the Everglades (data for calculation in Palmer and Mazzotti 2004, Brandt et al. 2010) the deep water refuge in LILA wetlands constituted 10–11% of the habitat area.
The aquatic flora and fauna were fairly consistent with what is common in the Everglades (Turner et al. 1999, Parkos et al. 2011). Wetland habitats are vegetated with common emergent and/or floating macrophytes (esp. Panicum hemitomon, Cladium jamaicense, Eleocharis cellulosa, Nymphaea odorata). Sloughs also contain submerged species, mainly Chara sp., Utricularia foliosa and U. purpurea. All wetland habitats are < 1 m deep except at the peak water levels in the late wet season (October) when the deep sloughs may exceed 1 m. The aquatic animal assemblages included crayfish (Procambarus fallax) and grass shrimp (Palaemonetes paludosus), libellulid dragonfly naiads (~5 common species) and other aquatic insects (e.g., Pelecoris femoralis) several species of small-bodied fishes (e.g., Gambusia holbrooki, Lucania goodei, Heterandria formosa) larger-bodied fishes like warmouth (Lepomis gulosus) dollar sunfish (L. marginatus) and Florida gar (Lepistosteus platyrhincus) as well as large predatory reptiles (Alligator mississippiensis) and amphibians (Rana grylio, Siren lacertina).
Fig. A1. Aerial photo of the Loxahatchee Impoundment Landscape Assessment (LILA) experimental wetlands in Palm Beach County, FL. Macrocosm labels and treatment designations were included for reference. The location of deep slough (DS) and shallow slough (SS) wetland habitats are indicated for M2. The other extensive habitats running the length of each macrocosm are higher elevation ridges that are inundated part of the year, but were extremely shallow or dry (mostly < 5 cm water depth) during our sampling seasons. Each deep slough also encircles two tree islands. The aircraft that took this photo was flying directly over the eastern side of the Arthur R. Marshall Loxahatchee National Wildlife Refuge. Deep ditches/ponds can be seen on the western end (bottom of photo) in each macrocosm. The alligator holes are too small to be visible in this photo. Photo Credit: South Florida Water Management District.
To measure mean large bodied fish (predator) catch rates (activity-densities) each of the four sampling seasons (two before manipulation, two after) we set and recovered (20–24 h sets) two pairs of fyke nets (set facing each other) of two types in the deeper slough of each wetland macrocosm. Two pairs of larger white fyke nets (0.9 × 1.2 m opening, 1 cm mesh) and two pairs of mini fyke nets (0.7 × 1 m opening, 3 mm mesh) were set facing each other with 15–20 m lead lines between them. The fyke nets were spread out and set in representative locations across each wetland in the same basic location (relative to shallow water and deeper ponds) in eachD wetland. In the shallower slough we set and recovered two pairs of black hoop nets (0.6 diam. opening, 1 cm mesh) one at the east end and one in the center. The hoop nets were set next to each other facing opposite directions (east or west) and wings (i.e., drift nets) were stretched laterally to block off the majority of the slough and force fish to either encounter a net opening or a shallow ridge habitat. Each wetland was fished for a total of 3 consecutive nights each sample season. A limited mark-recapture study suggested that the same fish were not being repeatedly caught (NJD, unpublished data). Because daily catch variation can be high (CV occasionally > 30% of sample mean) single catches reveal little the abundances in the wetlands; we collected data for three consecutive nights with the assumption that consistently high catches every day indicated large fishes were abundant. For logistical reasons the sampling had to be completed over two consecutive weeks (i.e., two wetlands were sampled each week).
B. Detailed methods and results from the fish removal/manipulation.
For fish removal and simulation of dry disturbance we drew the water down as low as possible with the culvert gates and gas powered pumps. When the two dried macrocosms were at their minimum we fished the ponds at the ends of each treatment wetland using multi-mesh gill nets (recovering them every few hours) and minnow traps. Live fish were released outside the LILA canal system. Dead fish collected during both phases of removal were buried. After three days of netting we applied rotenone to the perimeter ponds and ditches (still > 1 m deep). Rotenone was dripped into the ponds and ditches from a hose dragged behind a slowly moving airboat to achieve concentrations close to 3 ppm. Four low spots (i.e., simulated alligator holes) internal to each wetland retained 30–50 cm of water through the dry period and were not treated with rotenone, but were fished repeatedly with minnow traps. Visual inspection and minnow traps set in those spots indicated that they contained only smaller species of fishes (e.g., Jordanella floridae) and sustained a remnant population throughout the dry manipulation.
In total we physically removed approximately 24,000–25,000 fish (~647 kg wet mass, large and small fishes) combined from both wetlands with all techniques combined. The most abundant large-bodied fish species removed by gill nets and rotenone were warmouth (L. gulosus) and Florida gar (Lepisosteus platyrhincus). These removal estimates do not include the fish biomass removed by predatory wading birds and alligators during the drying period prior to the netting and rotenone application.
C. Central Everglades crayfish and small fish sample sites and hydrological covariates
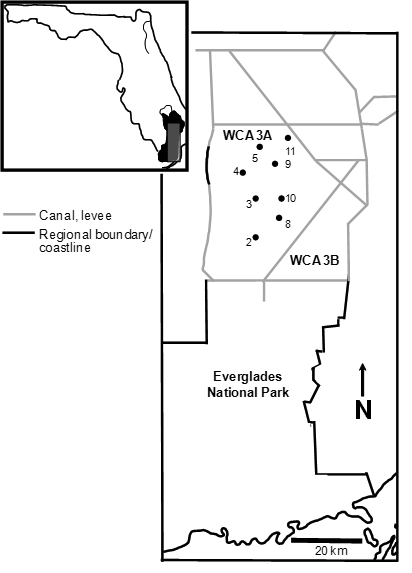
Throw trap sample sites were located throughout the central Everglades (Water Conservation Area 3A south of Alligator Alley; 25°59’34” N, 80°41’39” W; Fig. A2) in areas with low soil phosphorus and deep (> 1 m) organic soil (Osborne et al. 2011). The sites contained a mixture of sawgrass ridge (15–20 cm elevated soils dominated by Cladium jamaicense) and deeper slough (dominated by Nymphaea odorata or Eleocharis spp.) habitats, but throw trap locations were randomly located in the slough habitat only. Sawgrass ridges are logistically challenging to sample, but sampling of both habitats in adjacent areas suggest that crayfish densities are similar when both habitats are inundated (van der Heiden 2012). Sites were separated by > 4 km and were assumed to have effectively independent populations of the fish and crayfish (Ruetz et al. 2005). Sites 3 and 4 in Fig. A2 were included starting in 2010 and only sampled over the last three water years.
Water surface data for 20–30 sampling points (using GPS coordinates) for each of our sites (i.e., over several seasons) was downloaded from the EDEN water surfaces model that available online (V2. http://sofia.usgs.gov/eden/models/watersurfacemod_download.php; U. S. Geological Survey 2006). Using our own depth measurements from the random points (i.e., throw traps) and the associated water surfaces from the coordinates on the same we developed site-specific offsets relating surface level data (EDEN data are in cm relative to a universal standard) to average water depths (cm from sediment surface) in the sloughs. Hydrographs were then generated for each of our sites. From the hydrographs we generated measurements of the past hydrological conditions leading up to each of our sampling events. Hydrological variation including the average depth over the past 360 days (avg360) from the time of the trapping and the length of time a site was dry (depth < 1 cm) in the previous dry season (LD; an integer bounded by zero for sites constantly inundated). The two measures of past conditions are correlated (r = -0.69), but not identical, the avg360 covariates are running averages (changing within a water year), while LD is static for the entire water-year. Furthermore, the avg360 covariate incorporates high water depths as well as low water depths conditions. During the strong drought of 2011 the Everglades experienced extreme seasonal drying and even our wettest slough sites in WCA 3A dried to the sediment surface and several of the water gages in the Everglades also failed (i.e., water dropped below gage limits) during that period of time making the EDEN surface water data unreliable. For that period of time (spring/summer 2011) I used data from the nearest water gages that continued to work and created hydrographs using gage depths to predict slough water depths (cm) (simple linear regressions, similar to Ruetz et al. 2005, Dorn and Trexler 2007).
We included models with and without interactions between the hydro-covariates and seasons because the predominant mortality factors leading up to observed densities as well as population phenology are expected to vary between seasons and could create lagged density responses. For example, wading bird predation and direct effects of drying kill small fish and crayfish in the 1–2 months just prior to the summer sampling, but wading birds are largely absent in the summer and aquatic predators are expected to be more important sources of mortality throughout the wet season leading up to winter sampling. Wading bird predation in the dry season and desiccation-related stress may mute fish or crayfish densities in the summer, but the suppression of predatory fishes by the same dry disturbance may enhance recruitment success throughout the next wet season. We expected stronger hydrological correlations for crayfish for the winter (January) samples because our overall hypothesis has explicitly to do with effects of aquatic predators (e.g., fish) on crayfish recruitment occurring during the wet season. Furthermore, we expect juvenile crayfish may be present in the summer regardless of the recent hydrologic conditions. For small fishes we expected that the effects of recent drying might be strongest in the wet season (August) shortly after re-flooding (see Ruetz et al. 2005).
Fig. A2. Map of throw-trap sampling sites in the central Everglades (Water Conservation Area 3A). Each site was 25 ha (500 × 500 m) in area and was sampled with 5 throw traps in the summer (August) and winter (January). Sites 3 and 4 have were sampled for three full water years (2010–2012), the others were sampled from winter 2005 through winter 2013 (data from 8 water years). All sample site locations on the map are approximate.
Literature Cited
Aich, S., T. W. Dreschel, E. A. Cline, and F. H. Sklar. 2011. The development of a geographic information system (GIS) to document research in an Everglades physical model. Journal of Environmental Science and Engineering 5:289–302.
Brandt, L. A., M. R. Campbell, and F. J. Mazzotti. 2010. Spatial distribution of alligator holes in the central Everglades. Southeastern Naturalist 9:487–496.
Dorn, N. J. and J. C. Trexler. 2007. Crayfish assemblage shifts in a large drought-prone wetland: the roles of hydrology and competition. Freshwater Biology 52:2399–2411.
Osborne, T. Z., S. Newman, D. J. Scheidt, P. I. Kalla, G. L. Bruland, M. J. Cohen, L. J. Scinto and L. R. Ellis. 2011. Landscape patterns of significant soil nutrients and contaminants in the Greater Everglades ecosystem: past, present, and future. Critical Reviews in Environmental Science and Technology 41:121–148.
Palmer, M. L. and F. J. Mazzotti. 2004. Structure of Everglades alligator holes. Wetlands 24:115–124.
Parkos, J. J., C. R. Ruetz, and J. C. Trexler. 2011. Disturbance regime and limits on benefits of refuge use for fishes in a fluctuating hydroscape. Oikos 120:1519–1530.
Ruetz, C. R., J. C. Trexler, F. Jordan, W. F. Loftus, and S. A. Perry. 2005. Population dynamics of wetland fishes: spatio-temporal patterns synchronized by hydrologic disturbance? Journal of Animal Ecology 74:322–332.
U.S. Geological Survey. 2006. The Everglades depth estimation network (EDEN) for support of ecological and biological assessments. Fact Sheet 2006-3087, U.S. Department of Interior, Geological Survey, Ft. Lauderdale, Florida. http://sofia.usgs.gov/publications/fs/2006-3087/index.html, accessed 10 October 2013.
Turner, A. M., J. C. Trexler, C. F. Jordan, S. J. Slack, P. Geddes, J. H. Chick, and W. F. Loftus. 1999. Targeting ecosystem features for conservation: standing crops in the Florida Everglades. Conservation Biology 13:898–911.