Ecological Archives E096-250-A5
E. F. LoPresti, I. S. Pearse, and G. K. Charles. 2015. The siren song of a sticky plant: Columbines provision mutualist arthropods by attracting and killing passerby insects. Ecology 96:28622869. http://dx.doi.org/10.1890/15-0342.1
Appendix E. Exudate removal experiment.
We originally planned this experiment to compare to the carrion-removal experiment to compare any direct effects of exudates (direct deterrance of both herbivores and predators); the populations ended up being extremely different in phenology and morphology and a direct comparison was impossible. This population suffered the effects of the 2014 drought; this seep dried up in July and most plants senesced quickly. Because the manipulation also removed carrion, we feel these results are important to present, as it is a replication of sorts.
Methods
To determine whether carrion mediated an indirect defense with predators, we conducted an exudate- and carrion-removal experiment on an isolated population of columbine (38.852290N, 122.404978W). On 7 July 2014, we marked 70 plants, alternating between two treatments. The first treatment was an exudate removal, where we gently wiped the entire peduncle and pedicels with a soft damp sponge, which reduces stickiness by removing some exudates and removes nearly all carrion (pers. obs.). The second group was a control, which we handled as in the manipulated group, but did not remove any carrion. I repeatedly applied these at least biweekly (14 July, 20 July, 1 Aug, 5 Aug, 17 Aug, 25 Aug, 31 Aug, 8 Sept, and 18 Sept, treatments not reapplied the last two dates) treatments to the plants; each time also recording the number of carrion (except at set-up) and predators, and the number and status (damaged/undamaged) of all reproductive structures (flower buds/flowers/fruit) on the plant.
Results
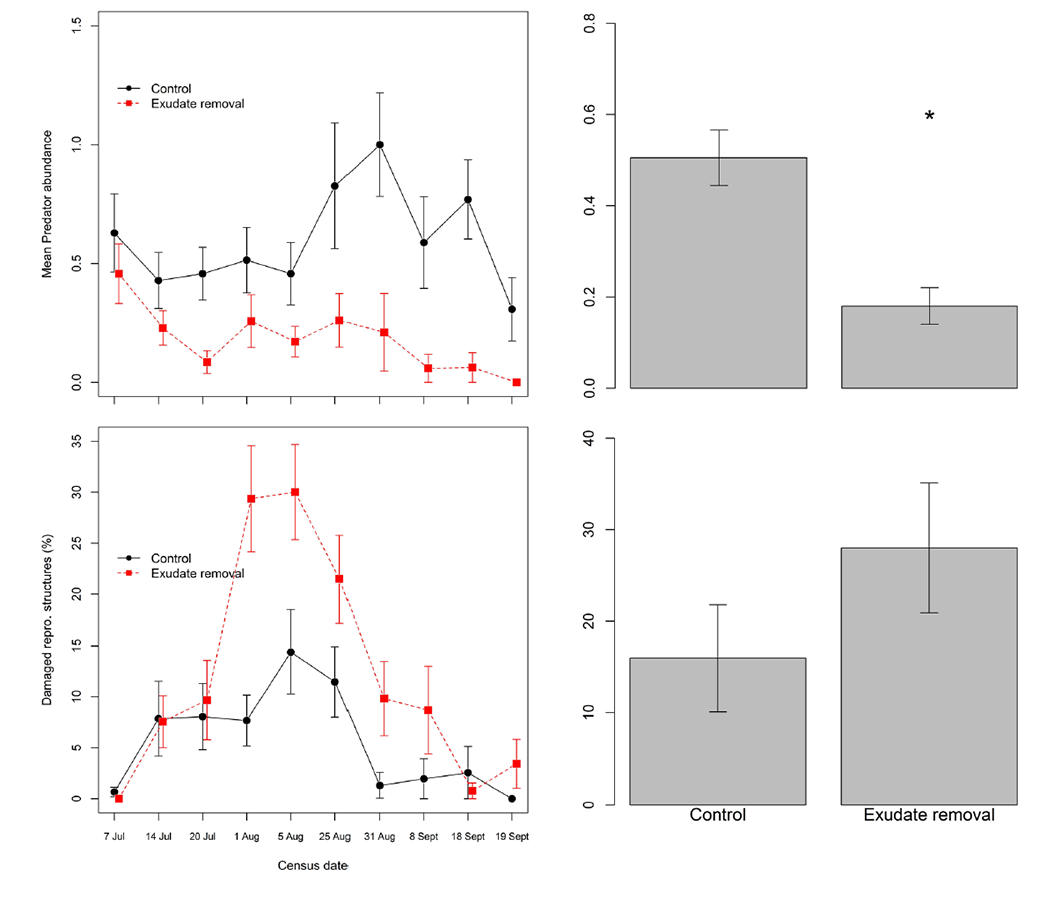
Mean predators encountered per check per plant was 183% higher in control than on carrion-removal plants (control 0.51 + 0.06, carrion-removed: 0.18 + 0.04: Fig. E1-1), (negative binomial mixed model on overall means, χ² = 13.54, p < 0.0001). Including the entire season (Fig. E1-2), a mixed model including treatment and date fit the predator data significantly better than a null model only including date (χ² =14.94, df = 1, p < 0.001), and the treatment effect was highly significant (t = -3.739, p = 0.002). Reproductive structures (flowers, flower buds, and fruits) of carrion removed plants were more significantly likely to sustain damage (STATS) than those of control plants, especially in the latter half of the season (Fig. E1-3). Including the entire season (Fig. E1-3), a mixed model including treatment and date fit the predator data significantly better than a null model only including date (χ² =39.65, df = 1, p < 0.0001), and the treatment effect was highly significant (t = -6.091, p < 0.0001).
Most plants dried up before any seed set could occur (control: 17/35, exudate removal 18/35); nearly all of the mortality occurred at the lowest part of the seep, which dried up most quickly. As a result, seed data does not reflect observed damage (see Appendix C) and is further limited by sample size and excessive zeroes. Excluding the first collection date, controls averaged nearly 100% higher fruit set, a marginally significant result (control 1.0 + 0.2, exudate removal 0.5 + 0.1, two-tailed t test, p = 0.08); higher total seeds (c: 67.0 + 17.7, er: 31.7 + 7.8, two-tailed t test, p = 0.07); higher seeds per centimeter of stem (c: 0.63 + 0.16, er: 0.32 + 0.08, two-tailed t test, p = 0.08) but similar seeds per fruit (c: 64.2 + 6.4, er: 60.3 + 11.0, two-tailed t test p = 0.77). Excluding the first 24 plants (12/treatment), which were in the lowest reach of the seep and noted as drying on 20 July (less than two weeks after initiation of experiment), the results became more robust (and meet statistical assumptions better) despite the smaller sample size: control plants had higher fruit set (c: 1.0 + .28, er: 0.8 + 0.1, two tailed t test, p = 0.03), higher seed set (c: 102.0 + 19.4, er: 47.8 + 8.4, two-tailed t test, p = 0.05), higher seeds per centimeter of stem (c: 0.97 + 0.22, er: 0.49 + 0.11, two-tailed t test, p = 0.05) but similar seeds per fruit (c: 64.2 + 0.66, er: 63.6 + 11.2, two-tailed t test, p = 0.97 ) perhaps because of lower resources for caterpillars on these much smaller plants.
Interpretation
The removal of carrion in this experiment, represents a partial replication and parallels the results of the carrion-removal experiment and Krimmel and Pearse (2013). While it would be tempting to conclude increased effect size of predator abundance was a result of the removal of both carrion and exudates, we caution any comparison of the two populations. As stated earlier, this population had far smaller plants and heavy deer herbivory. The deer herbivory did not influence the damage totals, since deer consume entire peduncles and pedicels, and damage analyzed was damaged and intact fruit. Any effect of the exudate itself will need to be tested in a factorial experiment with carrion removal within a single population.
Fig. E1. Upper left: Predator abundance on columbines with experimentally removed exudates (and carrion) and controls over the growing season. Upper right: mean arthropods per check, averaged over each plant over the season. Lower left: mean percentage reproductive structures damaged on columbines with experimentally removed exudates (and carrion) and controls over the growing season. Lower right: mean percentage damaged reproductive structures averaged over each plant over the season. All error bars are means +/- SE.
Literature Cited
Krimmel, B. A., and I. S. Pearse. 2013. Sticky plant traps insects to enhance indirect defence. Ecology Letters 16:219–24.