Ecological Archives E096-115-A5
Steve J. Kroiss and Janneke HilleRisLambers. 2015. Recruitment limitation of long-lived conifers: implications for climate change responses. Ecology 96:12861297. http://dx.doi.org/10.1890/14-0595.1
Appendix E. Potential impact of seed viability on results and conclusions.
We did not experimentally assess seed viability in this study. As such, we were unable to formally assess how seed viability varied across species' ranges or between masting and non-masting years. We wished to verify that such variation in seed viability (if present) would not have influenced our qualitative decisions about the importance of seed and microsite availability across species ranges and years. We used several approaches to tackle this issue.
First, we explored how seed viability varies across species ranges and years based on previously collected data from our study system. A small-scale germination trial by Ailene Ettinger (unpublished) in our lab examined seed viability for three of our study species (Abies amabilis, Tsuga heterophylla, and Tsuga mertensiana) using seeds from masting (2010) and non-masting years (2011), collected from multiple elevations (680–1620 m) across the range of each of our species: 4 sites for Abies amabilis, 2 sites for Tsuga heterophylla, and 3 sites for Tsuga mertensiana. A total of 600–940 total seeds were collected for each species. Seeds were germinated in trays of potting mix in a greenhouse. To examine the effect of elevation and year on germination rates, we fit generalized linear mixed effects models with random effects of stand to the data.
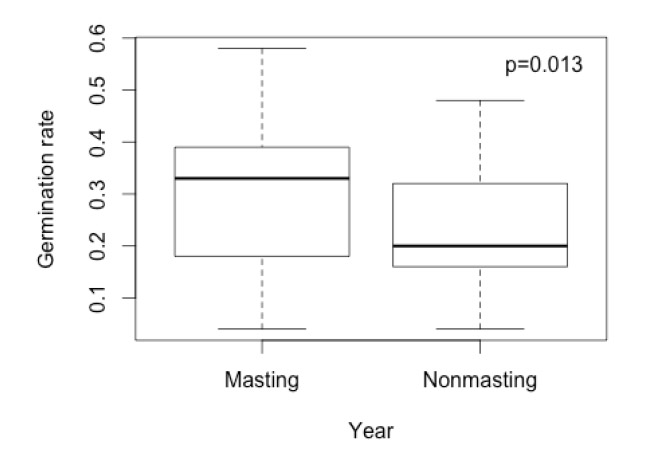
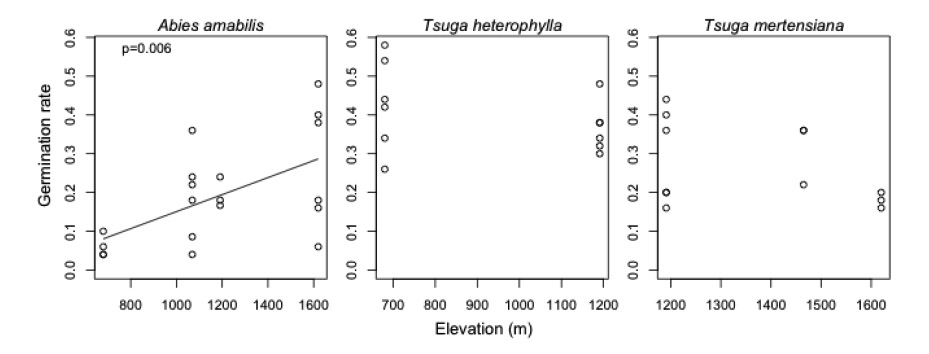
Analysis of these small-scale germination trials demonstrated that only one species (Abies amabilis) showed any evidence of variation in seed viability (i.e., germination) across elevation (Fig. E1). However, seed viability did vary temporally (Fig. E2) for all species. Specifically, germination was 1.24 times higher in masting years (ranging from 0.44 to 0.58 across species) than in non-masting years (ranging from 0.04 to 0.26 across species). Thus, there is some evidence to suggest that seed viability may vary across at least some species' ranges and between masting and nonmasting years. Below, we discuss the potential effects of spatial and temporal variation in seed viability on each of our major conclusions.
Our first conclusion, that seed availability is related to conspecific tree basal area and varies annually, is unlikely to be altered by spatial or temporal variation in seed viability. First, structured variation in seed viability across species' ranges is only present for 1 of 3 species in the germination trial. Even when present, such a relationship may alter the amount of noise in the relationship between basal area and seed availability, but would not remove it entirely, because basal area is not linearly related to elevation for any of our focal species. Higher seed viability in masting years (as found in the germination trial) implies that year-to-year variation in seed availability is even greater than what we found (making our conclusion about high year to year variability in seed production conservative).
Similarly, structured variation in viability across species ranges is unlikely to alter our second conclusion, that seed availability tends to decline towards range edges (with decreasing conspecific basal area). Seed viability is sometimes thought to decline towards range edges (i.e., peak in the core of range) – if this was the case, our conclusions about low seed availability at range edges would be conservative. If seed viability was highest at range edges, however, seed availability may not decline towards range edges as we concluded. This pattern in seed viability is unlikely (and not what was found for either Abies amabilis or Tsuga mertensiana, in the small-scale germination trial conducted by Ailene Ettinger), but we explored the implications of high seed viability at range edges on our conclusions anyway. Specifically, we used simulations to determine how much seed viability would need to increase at range edges to affect the significance of our observed relationships between seed availability and range position. We found that the relationship between seed availability and range position would only become nonsignificant if seed viability were about 25 times higher at the upper range edge than the core for Tsuga heterophylla, which seems unlikely given the existing range of seed viability found for this as well as the other three species (Fig E1).
Structured variation in seed viability across species' ranges could also alter our conclusion that fecundity (seeds per basal area) did not vary across species ranges. To examine this issue, we ran simulations to determine how much viability would need to decline at the range edge in order for us to observe a significant relationship between fecundity and range position. These simulations demonstrated that significant differences in fecundity (viable seeds / basal area) would only be observed if viability at range edges were reduced by 99% for Abies amabilis, 50% for Tsuga heterophylla, and 59% for Tsuga mertensiana compared to the range core. Similarly large increases in seed viability are required to change patterns in fecundity across ranges. While structured variation in seed viability across species ranges is possible, the magnitude and pattern of existing variation (Fig. E1) from germination trials does not imply such structured variation qualitatively influenced our conclusions about fecundity not declining at range edges.
Variation in seed viability across species' ranges could theoretically influence the relationship between microsite variables and germination, but we believe our qualitative conclusions about the identity and effects of microsite variables on germination (for different species) are robust for several reasons. First, our germination models included random effects of year and stand. Since all microsite variables were measured at the seedling plot level, and seed viability is unlikely to systematically vary within stands (since seedling plots are close together), these random effects should accommodate stand-to-stand and year-to-year variation in seed viability.
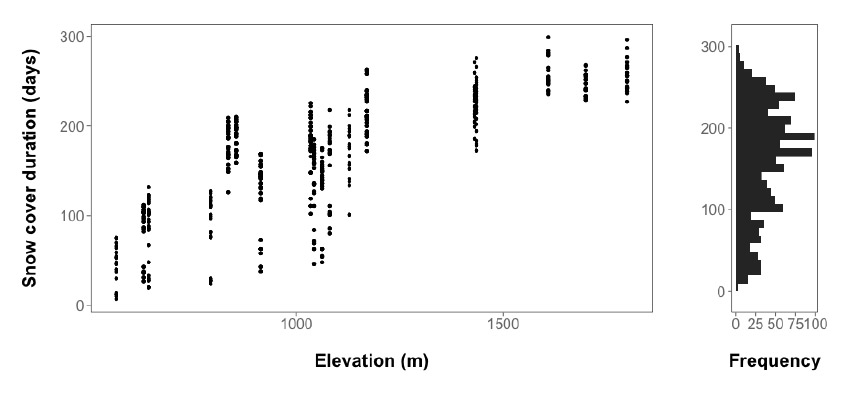
However, seed viability that varies with elevation could result in a spurious relationship between a microsite variable and germination (calculated as germinants / all seeds), if that microsite variable also varies with elevation. Only one explanatory variable, snow disappearance date, varied predictably with elevation (and thus range position; Fig. E3). However, we believe the effects of snow duration on germination we found in our statistical models are robust to any elevational trends in seed viability (if they indeed occur), for several reasons. First, seed viability would have to vary strongly with elevation, but differently among species, for us to have observed species-specific differences in responses to snow disappearance date, which seems unlikely. Seed germination trials did suggest seed viability increases with increasing elevation for Abies amabilis alone (and thus, potentially with greater snowpack duration – Fig. E1), but the relationship between snowpack duration and germination for this species was opposite of the direction that seed viability-elevation patterns would drive it (suggesting that our results for this species may be conservative). Finally, there was considerable variation in snowpack duration within each forest stand, allowing us to disentangle snow duration effects on seed germination when seed inputs have a common viability within one stand (Fig. E3).
Finally, variation in seed viability between masting and nonmasting years is unlikely to alter our fourth conclusion, that years of high seed production were generally correlated among species, but not correlated with the availability of suitable microsites. To examine this issue, we simulated the potential effects of annual variation in seed viability by recalculating seed availability and germination rates based on the maximum observed seed viability differences from germination trials. Specifically, we recalculated seed availability (# viable seeds) in nonmasting and masting years using the minimum (0.04) and maximum (0.58) species-specific seed viability values from the germination trials respectively. Correlations between seed availability and germination within species were qualitatively unchanged (i.e., insignificant) under this scenario. Specifically, only one species demonstrated significant correlations between seed availability and germination in original correlations, which was the case under these simulations, although identity of the species exhibiting this pattern shifted from Tsuga heterophylla to Abies amabilis. Correlations of high seed production years among species were also qualitatively unchanged when modifying seed availability according to seed viability in masting vs. non-masting years. In fact, simulations suggested that our results are conservative, since the number of significant pairwise correlations among species increased from four out of eight to seven out of eight.
Fig. E1. Relationships between germination rates and seed source elevation from small-scale greenhouse germination trials. Seed collections were made from the lower, core and upper ranges of Abies amabilis (range ~500–1800 m); from the core and upper ranges of Tsuga heterophylla (range below lowest elevation in park to 1500 m); and from the lower, core and upper ranges of Tsuga mertensiana (range ~900–1800 m). Significant relationships are shown with solid regression lines.
Fig. E2. Variation in germination rates between masting and nonmasting years based on small-scale greenhouse germination trials.
Fig. E3. The left panel shows that snow cover duration increases with elevation, but also exhibits considerable variation within each of the 18 forest stands in the study area. The right panel shows a histogram of snow duration values.