Ecological Archives E096-045-A2
Grace F. Chen and Douglas W. Schemske. 2015. Ecological differentiation and local adaptation in two sister species of Neotropical Costus (Costaceae). Ecology 96:440449. http://dx.doi.org/10.1890/14-0428.1
Appendix B. Detailed descriptions of the transplant sites in central Panama.
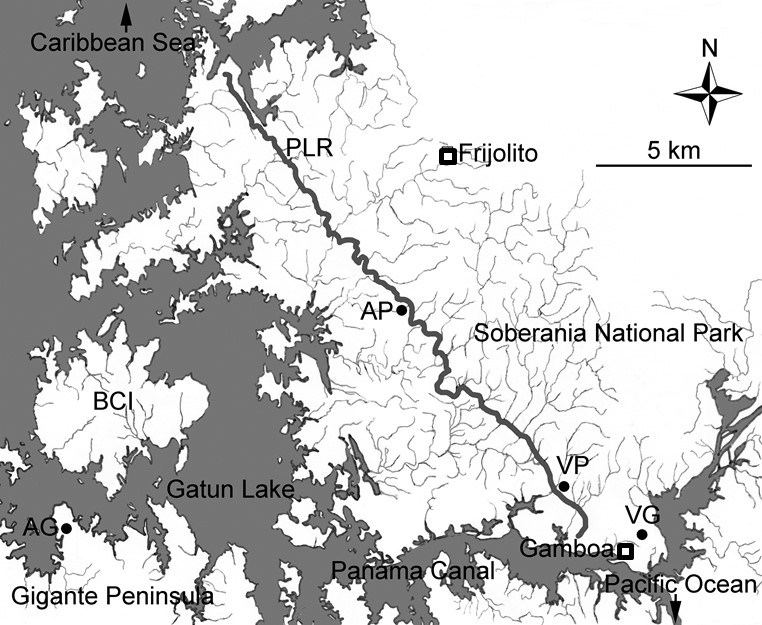
Fig. B1. Map of study sites in Central Panama. Pipeline Road (PLR) is marked by the thick solid line. Precipitation data were collected at the Frijolito and Gamboa weather stations marked with open squares. Closed circles indicate the location of the reciprocal transplant sites: C. allenii sites on Pipeline Road (AP) and on the Gigante Peninsula (AG), and C. villosissimus sites on Pipeline Road (VP) and in Gamboa (VG).
Across the Isthmus of Panama, a distance of 80 km, more rain falls on the Atlantic side (mean annual precipitation = 3234 mm at Cristobal) than on the Pacific side (mean annual precipitation = 1798 mm at Balboa Heights; Panama Canal Authority Meteorological and Hydrological Service weather station network). The field components of this study were conducted in Soberania National Park located along the Panama Canal approximately 25 km north of Panama City. The park encompasses 22,104 hectares, is located at the center of the Isthmus-wide rainfall gradient, and includes populations of both of the study species.
The spatial distribution of the two species was determined along a transect, Pipeline Road (PLR; Fig. B1), running through Soberania National Park (south – north). Pipeline Road spans the sharp precipitation gradient in the Canal region and provides access to natural habitats of C. allenii and C. villosissimus. Along the southern end of PLR, the road is wide and the canopy is open. As the road progresses northward, the canopy closes gradually and the road narrows until it becomes completely inaccessible. We studied the spatial distribution of the two species and made environmental measurements (soil moisture, light availability) in natural habitats along 17 km of PLR. Both species were found along this section, with C. allenii predominating in the northern stretch and C. villosissimus to the south.
For the reciprocal transplant experiments, we employed two transplant sites for each species (Fig. B1). For C. villosissimus, one site (VP) was near the center of the distribution of C. villosissimus on PLR, located between 1 km and 2 km from the entrance, and the other one (VG) was in Gamboa near the entrance of PLR. For C. allenii, one site (AP) is near the center of the distribution of C. allenii on PLR, located between 10 km and 11 km from the entrance, and the other one (AG) was on Gigante Peninsula in Barro Colorado Nature Monument (Fig. B1). With the exception of the C. allenii site on Gigante, all the other sites possess natural populations of the respective species. On Gigante, natural populations of C. allenii have not been located, probably because of previous disturbance. Nevertheless, the environmental conditions appeared similar to those of other C. allenii habitats. In the C. villosissimus site on PLR (VP), the forest is mostly second growth with semi-open canopy. At the other C. villosissimus site (VG), there is a forest-grassland transition that was a result of clearing of the forest. At both C. villosissimus sites, transplant transects were placed along the forest edge to mimic locations of natural populations. At the C. allenii site on PLR (AP), a ravine runs through the forest near the road and transplant transects were placed on the banks along the ravine. The other C. allenii site (AG) is located on the north margin of Gigante Peninsula and transplant transects were placed along the shore.
For seed transplants, 55 seeds from each species were transplanted into one C. allenii site (AP) and one C. villosissimus site (VP) on PLR in both 2007 and 2008 for a total of 440 seeds (55 seeds × 2 species × 2 sites × 2 years = 440 seeds). For clonal transplants, we planted 18 C. allenii and 18 C. villosissimus plants at the AG site and 13 C. allenii and 18 C. villosissimus plants at each of the C. villosissimus sites (VP and VG) in June 2006. In June 2007, we planted 13 C. allenii and 18 C. villosissimus plants at the AP site.