Ecological Archives A025-034-A4
Jeffrey F. Bromaghin, Trent L. McDonald, Ian Stirling, Andrew E. Derocher, Evan S. Richardson, Eric V. Regehr, David C. Douglas, George M. Durner, Todd Atwood, and Steven C. Amstrup. 2015. Polar bear population dynamics in the southern Beaufort Sea during a period of sea ice decline. Ecological Applications 25:634651. http://dx.doi.org/10.1890/14-1129.1
Appendix D. An analysis of movement data from southern Beaufort Sea polar bears instrumented with radio-collars.
INTRODUCTION
Movements of marked and unmarked individuals between capture occasions can be an important aspect of mark-recapture investigations because some types of movement have the potential to bias parameter estimates. For mobile species, emigration (movement to exit the study area) has received considerable attention in the mark–recapture literature. Emigration that is temporary and random, in the sense that each individual has a common probability of being within the study area, is not problematic. Under these conditions, recapture probability parameters can be thought of as the product of two probabilities, the probability of being within the study area and the conditional probability of capture given presence (Burnham 1993, Kendall et al. 1997, Schwarz and Stobo 1997). Estimates of survival remain unbiased and estimates of abundance include members of the population that have temporarily emigrated. Conversely, permanent emigration is more troublesome because it is indistinguishable from death (e.g., Nichols 2005). Permanent emigration reduces survival rate estimates because survival parameters incorporate both the probability of remaining in the study area and true survival of individuals that remain part of the population. For this reason, survival in Cormack-Jolly-Seber models is often termed "apparent survival". Estimates of recapture probabilities, from which abundance estimates are derived, apply to individuals that remain in the study area. A pulse of permanent emigration, perhaps in response to deteriorating ecological conditions, can appear as a temporary decline in survival and a permanent decline in abundance. Temporary emigration near the end of a mark-recapture investigation is difficult to distinguish from permanent emigration.
Movement patterns also can be problematic if individuals are only captured within a portion of the geographic range of a population. If the movement of individuals within the entire population range is random, the net effect is equivalent to random temporary emigration and parameter estimation is not adversely affected. However, patterns of individual movement could create non-random structure in recapture probabilities within the population. For example, some individuals may tend to prefer habitats within the capture region and other individuals may prefer habitats outside that region. Such a movement pattern is effectively equivalent to heterogeneity in recapture probabilities. The presence of un-modeled heterogeneity in recapture probabilities can bias estimates of recapture probabilities and abundance (Carothers 1973), though estimates of survival probabilities are more robust to heterogeneity (Carothers 1979, Abadi et al. 2013), but also see Fletcher et al. (2012). However, some patterns of Markovian movement between consecutive capture events can bias survival rate estimates (Kendall et al. 1997, Schaub et al. 2004).
METHODS
The potential influence of both emigration and availability within a restricted capture region are of interest in the mark-recapture investigation of southern Beaufort Sea polar bears. Deteriorating sea ice conditions (Fig. 2) may have altered the distribution of bears during the course of this investigation and are projected to do so in the future (Durner et al. 2009). In addition, polar bears can only be captured within the near-shore habitats within the range (~160 km) of capture helicopters (Fig. 1), and bears are known to roam further offshore (Amstrup et al. 2004). To detect potential changes in movement patterns that might have biased estimates, we analyzed movement data from radio collars deployed by the U. S. Geological Survey on female polar bears in the southern Beaufort Sea.
We analyzed location data from individual polar bears with radio collars between 1999 and 2010. Data were examined for evidence that bears had successfully removed their collars and locations subsequent to a suspected early removal were excised from the data. We restricted analyses to those animals for which location data were available within a 365 ± 30 day window from the date they were collared. Our primary interest was the location of bears during the spring capture season in the years of this investigation, so individuals with location data during the months of March to May each year from 2001 to 2010 were identified. These screening filters resulted in a data set containing 116 time series of spring locations from 71 unique bears.
Each bear in each year was classified as being either available or unavailable for capture based on its location data. We tested severalclassification criteria to ensure that results were not sensitive to the criteria used. Results are only reported for the analysis in which a bear was deemed available for capture if it had at least one location within 100km of the coast between -160º W and -141º W from 1 March to 31 May. Restricting the 100km distance criterion to 50km, or requiring an individual to have at least two locations within the study area separated by at least 14 days, had no influence on the results.
RESULTS
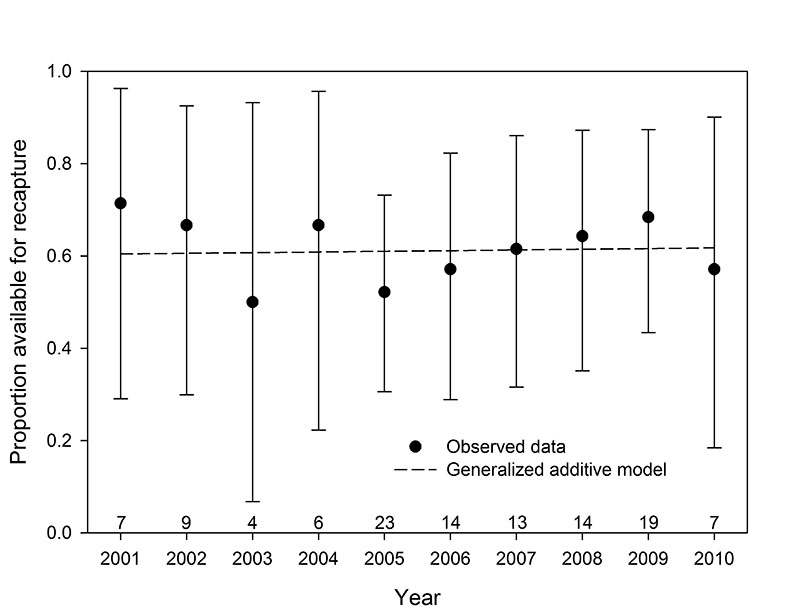
The proportions of radio-collared bears available for capture each year did not display a temporal trend (Fig. D1). The null hypothesis that the proportions were equal was tested using an exact binomial test (Agresti 2002), using 10,000 replications to estimate significance (Patefield 1981), and the proportions were not found to differ (p-value = 0.990). In addition, a generalized additive model (Wood 2006) with potential nonlinearity through time was used to approximate the proportion data. The estimated parameter for year was not significantly different than zero (p-value = 0.934), suggesting a straight line provided the best model fit to the data, which is consistent with the results of the exact binomial test.
Fig. D1. The proportion of radio-collared polar bears from the southern Beaufort Sea available for recapture in the study area, by year, with exact 95% confidence intervals and an estimated generalized additive model. The number of bears with location data each year is shown immediately above the horizontal axis.
The location data included 42 cases for which the availability of an individual within the Alaskan portion of the southern Beaufort Sea capture region in consecutive years could be determined (Fig. D2). The hypothesis of conditional independence between availability in consecutive years was tested using an exact Cochran-Mantel-Haenszel test (Agresti 2002). The test was significant (p value = 0.002), suggesting that availability within the capture region is not purely random between consecutive years.
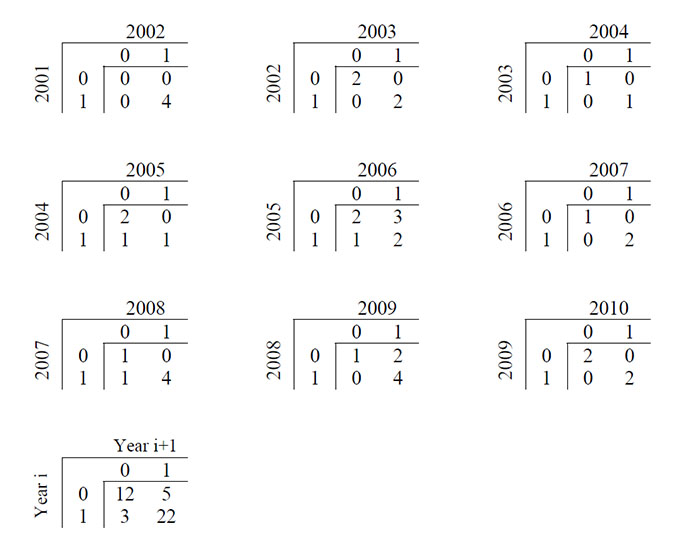
Fig. D2. Contingency tables cross-classifying the number of radio-collared polar bears available (1) or unavailable (0) for capture within the southern Beaufort Sea study area of Alaska in consecutive pairs of years from 2001 to 2010.
CONCLUSIONS
We found no evidence to suggest that the proportion of bears available within the capture region during the spring capture season has trended or otherwise changed during the study period (Fig. D1). Consequently, the reduced survival rates estimated for the middle portion of the study period (Fig. 3, 4) are unlikely to have been caused by a pulse in permanent emigration in response to unfavorable summer ice conditions (Fig. 2). The temporal consistency in the annual proportions reinforces the finding that a period of reduced survival during the middle portion of the study period led to a decline in abundance.
Although we found no reason to suspect animal movements within the study area could explain the temporal patterns in survival rate and abundance estimates, apparent non-randomness in movements of individuals between consecutive years was detected. Such Markovian temporary emigration has the potential to bias estimates of survival (Kendall et al. 1997, Schaub et al. 2004), as well as recapture probability estimates and therefore abundance estimates (Carothers 1973, Kendall et al. 1997, Schwarz and Stobo 1997). However, the observed pattern in movements between consecutive years (Fig. D2) is consistent with conditions under which the bias in survival probability estimates was low (Schaub et al. 2004). Consequently, although our tests corroborate a conclusion that the estimated decline in survival and abundance during the middle portion of the study period reflects actual population dynamics, the absolute magnitude of the estimates could incorporate some negative bias. We have no reason to suspect that such bias would differ substantially among years and potentially influence temporal trends in parameter estimates. In addition, the covariate Cap-procliv was designed to account for sources of heterogeneity in recapture probabilities and may partially abate the influence of any such bias (Appendix F).
Literature cited
Abadi, F., A. Botha, and R. Altwegg. 2013. Revisiting the effect of capture heterogeneity on survival estimates in capture-mark-recapture studies: Does it matter? PLoS One 8:e62636.
Agresti, A. 2002. Categorical data analysis, Second edition. John Wiley and Sons, Hoboken, New Jersey, USA.
Amstrup, S. C., T. L. McDonald, and G. M. Durner. 2004. Using satellite radiotelemetry data to delineate and manage wildlife populations. Wildlife Society Bulletin 32:661–679.
Burnham, K. P. 1993. A theory for the combined analysis of ring recovery and recapture data. Pages 199-213 in Lebreton, J.-D., and P. M. North, eds. Marked individuals in the study of bird populations. Birkhäuser Verlag, Basel, Switzerland.
Carothers, A. D. 1973. The effects of unequal catchability on Jolly-Seber estimates. Biometrics 29:79–100.
Carothers, A. D. 1979. Quantifying unequal catchability and its effect on survival estimates in an actual population. Journal of Animal Ecology 48:863–869.
Durner, G. M., D. C. Douglas, R. M. Nielson, S. C. Amstrup, T. L. McDonald, I. Stirling, M. Mauritzen, E. W. Born, Ø. Wiig, E. DeWeaver, M. C. Serreze, S. E. Belikov, M. M. Holland, J. Maslanik, J. Aars, D. A. Bailey, and A. E. Derocher. 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecological Monographs 79:25–58.
Fletcher, D., J. Lebreton, L. Marescot, M. Schaub, O. Gimenez, S. Dawson, and E. Slooten. 2012. Bias in estimation of adult survival and asymptotic population growth rate caused by undetected capture heterogeneity. Methods in Ecology and Evolution 3:206–216. 10.1111/j.2041-210X.2011.00137.x
Kendall, W. L., J. D. Nichols, and J. E. Hines. 1997. Estimating temporary emigration using capture-recapture data with Pollock's robust design. Ecology 78:563–578.
Nichols, J. D. 2005. Modern open-population capture-recapture models. Pages 88–123 in Amstrup, S. C., T. L. McDonald, and B. F. J. Manly, eds. Handbook of capture-recapture analysis. Princeton University Press, Princeton, New Jersey, USA.
Patefield, W. M. 1981. Algorithm AS159. An efficient method of generating r x c tables with given row and column totals. Applied Statistics 30: 91–97.
Schaub, M., O. Gimenez, B. R. Schmidt, and R. Pradel. 2004. Estimating survival and temporary emigration in the multistate capture-recapture framework. Ecology 85:2107–2113.
Schwarz, C. J., and W. T. Stobo. 1997. Estimating temporary migration using the robust design. Biometrics 53:178–184.
Wood, S. 2006. Generalized additive models: An introduction with R. Chapman and Hall/CRC, Boca Raton, Florida, USA.