INTRODUCTION
Spatial scale has been proposed as a unifying concept in ecology, and is intrinsic to the disciplines of biodiversity (Rosenzweig 1995), biogeography (MacNally et al. 2004), population biology (Fahrig 1992) and ecosystem ecology (Petersen et al. 1997). Indeed, scale is an important component of recent advances in ecology, such as metabolic theory (Marquet et al. 2004) and the unified neutral theory (Hubbell 2001). Nevertheless, the science of scale has been hampered by the paucity of scale-explicit data sets that simultaneously vary different components of scale.
There are numerous ways to decompose scale into its components, but here we consider grain and extent to be parameters of special interest (Allen and Hoekstra 1991). Grain is the size of the basic sampling unit, and extent is the size of the sampling universe. Both can be expressed either in terms of linear dimensions or area (or in three-dimensional systems, volume).
One crucial element for a study to be truly a scale study is objective placement of samples. While there are sometimes valid reasons for subjective sample unit locations, locating sample units subjectively to either minimize internal heterogeneity, maximize among-unit heterogeneity, maximizing ‘representativeness’, or stratification based on environmental criteria all interfere with our ability to detect a true signature of scale. Many have realized the importance of objective placement, and thus there are a wide variety of grid-based studies in the ecological literature (Heikkinen 1996, Bowman et al 2000, Palmer 2005, Bruun et al. 2003). Grids can easily be subsampled to allow a study of extent, but without spatially nested samples, there is limited ability to detect effects of grain.
Here, we present a scale-explicit data set of vascular plant species occurring in a grid consisting of nested square quadrats in the Oosting Natural Area in the Duke Forest of North Carolina, USA. This data set has already been used as a case study for questions of spatial scale and distance decay. Reed et al (1993) evaluated the scale dependence of vegetation/environment relationships. Palmer and White (1994) used the data to evaluate the scale-dependence of the species area relationship. Jonsson and Moen (1998) examined distance decay of species richness to evaluate optimum sampling designs. Richard (1998) examined the scale dependence of fern distributions in the Oosting preserve, and compared it to distributions in other regions. Wagner (2003) used the data to develop a method integrating variography and the variance test. Palmer et al. (2002) used the Oosting data in combination with other data to develop new methods for completing species inventories. Xi (2005) examined the effects of spatial scale on the interpretation of hurricane damage to forests.
It is our hope that making these data more widely available will encourage the development and evaluation of methods in spatial ecology.
|
|
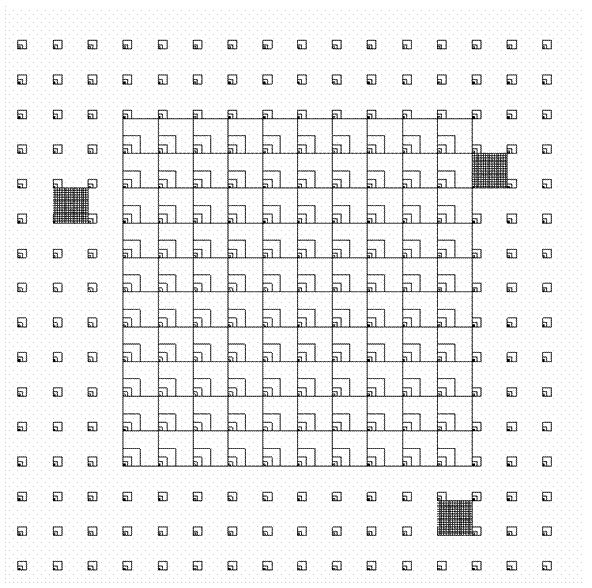
| FIG. 1. Five overlapping study grids in the Oosting Natural Area. The overall dimensions are 256 m × 256 m; details are given in the text and Tables 1 and 2. The five grids are: (1) All 256 Grid Cells, (2) Central 100 Grid Cells, (3) Fine-Scale #1, (4) Fine-Scale #2, and (5) Fine-Scale #3. Only plots of level 4 (1 m × 1 m) or higher (Table 1) are shown here. |
TABLE 1. Linear and area dimensions of square plots in the Oosting study grids (following Palmer and White 1994).
Linear dimensions (m) |
Area (m2) |
|
1 |
0.125 × 0.125 |
0.015625 |
2 |
0.25 × 0.25 |
0.0625 |
3 |
0.5 × 0.5 |
0.25 |
4 |
1.0 × 1.0 |
1 |
5 |
2.0 × 2.0 |
4 |
6 |
4.0 × 4.0 |
16 |
7 |
8.0 × 8.0 |
64 |
8 |
16.0 × 16.0 |
256 |
TABLE 2. The five study grids used for the vascular plant presence data. The (0,0) point is considered to be the southwest corner of the entire grid. See Table 1 for the scales indicated by ‘level’. Maximum extent is presented in cardinal directions only, multiply by square root of 2 to get maximum diagonal extent. Soils data and tree mapping were performed for the entire 256 m × 256 m study site. Grids 3, 4, and 5 have identical dimensions.
| Grid number | Grid name |
Easting of origin (m) |
Northing of origin (m) |
Levels represented |
Minimum extent (m) |
Maximum extent (m) |
1 |
All 256 |
0 |
0 |
1,2,3,4,5,6 |
16 |
256 |
2 |
Central 100 |
48 |
48 |
1,2,3,4,5,6,7,8 |
16 |
160 |
3 |
Fine-Scale #1 |
16 |
160 |
1,2,3,4 |
1 |
16 |
4 |
Fine-Scale #2 |
192 |
16 |
1,2,3,4 |
1 |
16 |
5 |
Fine Scale #3 |
208 |
176 |
1,2,3,4 |
1 |
16 |
METADATA CLASS I. DATA SET DESCRIPTORS
A. Data set identity:
Title: Vascular plants in a multiscale grid of plots in the Oosting Natural History Area, North Carolina
B. Data set identification code
Suggested Data Set Identity Code: oosting_Grid
C. Data set description
Principal Investigator: Michael W. Palmer, Department of Botany, Oklahoma State University, Oklahoma USA
Peter S. White, North Carolina Botanical Garden and the University of North Carolina, Chapel Hill, North Carolina, USA
Robert K. Peet, University of North Carolina, Chapel Hill, North Carolina, USA.Abstract:
We present a data set collected in 1989 of vascular plant occurrences in overlapping grids of nested plots in the Oosting Natural Area of the Duke Forest, Orange County, North Carolina, USA. The purpose of these data is to allow the study of vascular plant biodiversity at multiple spatial scales, in terms of both grain and extent. There are eight distinct plot sizes (grains), with size four times that of the next smallest, from 0.015625 m2 to 256 m2. Larger grains are possible by aggregating adjacent plots. Extents (linear distances between plots) range from 1 m to 362 m. In addition, trees were mapped over the entire 256 m × 256 m study site in 1990, and again in 1998 to assess the effects of Hurricane Fran in September 1996. Tree species, diameters, and damage descriptors (1998 only) were also recorded. Soils were gathered from the entire grid in 1990 and analyzed for nutrients, pH, and organic matter. These data have already been used to assess scale dependence of plant diversity, to evaluate new methods for the study of spatial scale, and to study the importance of spatial scale on the interpretation of hurricane damage to forests. They are presented as a case study for future scale research.
D. Key words: distance decay; hurricane; North Carolina; soil nutrients; spatial scale; species–area relationships; tree mapping; vascular plants.
CLASS II. RESEARCH ORIGIN DESCRIPTORS
A. Overall project description
Identity: A multiscale plant biodiversity study of the Oosting Natural History Area
Originators: M. W. Palmer, P. S. White, R. K. Peet, R. A. Reed, W. Xi.
Period of Study: multiscale vascular plant data from May and June, 1989; tree mapping from 1990 and 1998; soil data from 1990.
Objectives:The objectives of this study are to evaluate species–area relationships and patterns of distance decay of vascular plant distributions, in an objective, scale-dependent manner.
Abstract: same as above.
Source(s) of funding: North Carolina Botanical Garden; Oklahoma State University College of Arts and Sciences; NSF grants BSR-8905926 and DEB97-07551 to RKP.
B. Specific subproject description
Site description:
The Oosting Natural Area (ca. 65 ha in size), is located at 79.057W, 35.976N, and is part of the Duke Forest in Orange County, North Carolina. Elevations in this area range from about 100 to 150 m above sea level. This second-growth Piedmont hardwood forest is dominated by Liriodendron tulipifera (tulip poplar), Liquidambar styraciflua, several species of Quercus (primarily Q. rubra, Q. alba), several Carya species, (primarily C. tomentosa), and two species of Pinus (taeda, echinata)(see Appendix 1 of Reed et al. 1993). This community is equivalent to the Dry-Mesic Eutrophic Forest of Peet and Christensen (1980), the Basic Oak-Hickory Forest of Schafale and Weakley (1990), and the Quercus rubra-Quercus alba-Carya glabra/Geranium maculatum Forest of the U.S. National Vegetation Classification (NatureServe 2007). The soils that support such vegetation are primarily of the Iredell series, derive from intrusive mafic rock, have higher pH and available calcium than is typical for the region, and have subsoils with significant content of shrink-swell clays. The mean pH of 6.0 and mean available Ca of 1395 ppm obtained from the 289 soils samples (see Appendix 2 of Reed et al. 1993) greatly exceed the median values of around 4.4 and 160 respectively obtained by Peet and Christensen (1980) in their survey of Duke Forest vegetation . However, the range of values found in this study does fall within the range found in their study.
The study area has been owned and protected by Duke University since the mid-1940s and is managed for conservation and non-manipulative research. This area was included in the North Carolina Registry of Natural Heritage Area in 2004. The relatively even size distribution of the dominant trees, together with the abundance of such typically successional species as Liriodendron and Liquidambar, suggests that disturbance influenced tree establishment, probably in the late 1800s. With the exception that a considerable population of small (
3 – 10 cm stump diameter) Juniperus virginiana was cut, probably between 1930 and 1950, there is no evidence of significant disturbance between 1900 and Hurricane Fran in 1996.
The Duke Forest flora consists of 978 vascular plant species (Palmer 1990). Referenced voucher specimens are deposited in the Duke Herbarium (herbarium code: DUKE), Duke University, Durham, North Carolina, USA).Research Methods:
We placed a complex of five overlapping sampling grids (Fig. 1; Table 2) randomly within the Oosting Natural Area, subject only to the constraint that the 256 × 256 study site fit entirely within the Natural Area. The five grids are (1) All 256 Grid Cells, (2) Central 100 Grid Cells, (3) Fine-Scale #1, (4) Fine-Scale #2, and (5) Fine-Scale #3, and are further described in Table 2. Grids 3, 4, and 5 have identical dimensions; thus there are three types of grid. A nested series of square quadrats of levels 1 through 6 (with dimensions given in Table 1) is located in the southwestern corner of each grid cell; the southwestern corner of each quadrat for a grid cell is shared. Larger quadrats (including levels 1 through 8) were established in the central 100 grid cells. We randomly selected three grid cells (the fine grids in Fig. 1) for more intensive study; in these, every square meter was sampled using nested quadrats of levels 1 through 4. We thus present the data from a total of five grids (Table 2), although in some cases the actual quadrats used are coincident.
The grid was marked using stakes constructed of PVC pipes; a visit by Jason Fridley in 2006 (personal communication) indicated that the majority of the stakes are still in place. Thus, a resurvey of the grid is possible. Upon establishment, the relative elevation of the corners of the grid cells was assessed using a surveyor’s transit and rod.
Between May and June 1989, we recorded the presence of vascular plant species in each of the nested quadrats. Because they are nested, the list of species in a quadrat of a given level must be a subset of the species in the next higher level. Individuals not immediately identifiable in the field were marked for later identification; in nine cases these could not be found upon revisiting. However, these individuals are not likely to belong to any of the identified species in the data matrix and thus would probably not lead to errors in assessing diversity.
In the summer of 1990, and again in 1998 (to assess the impact of Hurricane Fran in 1996; Xi 2005) we identified and mapped all trees >2cm diameter at breast height to the nearest 0.1m, and recorded diameter to the nearest 1mm, within the entire 256m × 256m study site. We also assessed in 1998 the nature of the hurricane damage to individual stems. During the 1998 surveys, in addition to continuing measurement of vitality, diameter, and survival, we quantified hurricane damage status for each individual stem in the plot with stem damage codes. The stem damage codes we used were uprooting (no such damage, partial uprooting, complete uprooting), breakage (no such damage, <10%, 10–35%, 35–90%, and >90% canopy loss), leaning (no such damage, lean over 10%, supported by another tree, down on ground), and leaned on (no such damage, upright, supporting a tree, bent or leaning, pinned). The azimuth of each fallen bole was measured by compass.
Soil samples were collected at the corners (289) of all 256 grid cells in 1990 from the top 10 cm of mineral soil after removal of the litter layer. Soils were dried and sieved to 2 mm before shipping to Brookside Laboratories (New Knoxville, Ohio), where they were analyzed for total exchange capacity, pH, percent organic matter, bulk density, and (using a Mehlich 3 extractant; see Mehlich 1984) available sulfur, phosphorus calcium, magnesium, potassium, sodium, boron, iron, manganese, copper, zinc, and aluminum. Although small-scale spatial variation in chemistry of soils derived from igneous rocks can be considerable (Palmer and Dixon 1990), the fact that soil chemistry as assessed from a single soil sample in a 1000-m2 plot has been shown to be a strong predictor of vegetation composition (e.g., Peet and Christensen 1980, Reed et al. 1993, Newell and Peet 1998) in this region gives us confidence that our 289 soil samples were sufficient to adequately characterize the site.
Nomenclature follows Radford et al. (1968) but we also report synonyms according to Kartesz (1999).
Project Personnel: M. W. Palmer was responsible for establishing the grid and gathering all data on the grid, but was assisted at times by K. Doyle, S. D. McAlister, I. L. Palmer, S. S. Palmer, R. A. Reed, M. Robinson, and K. Wolf. R. A. Reed, W. Xi, M. Cassey, K. Jacobs, J. Shepherd, C. Rader, L. Meissner, R. White, R. McDonald, and R. K. Peet gathered the tree and soil data.
CLASS III. DATA SET STATUS AND ACCESSIBILITY
A. Status
Latest Update: July 2007 for the final format of all files.
Latest Archive date:July 10, 2007
Metadata status: Metadata are complete for this period and are stored with the data (see B. below).
Data verification: Understory data were visually proofed by M.W. Palmer and S. D. McAlister to match written data sheet records with the computer files. Soil and tree data were visually checked by R. A. Reed and W. Xi, and examined for logical inconsistencies and unlikely values by R. K. Peet, J. Ott, and M. F. Boyle.
B. Accessibility
Storage location and medium: All digital data exist on first author’s personal computer in ASCII format.
Contact person: Michael W. Palmer, Department of Botany, Oklahoma State University, Stillwater Oklahoma 74078 USA; tel 405-744-7717; fax 405-744-7074; [email protected].
Copyright restrictions: None
Proprietary restrictions: None
Costs: None
CLASS IV. DATA STRUCTURAL DESCRIPTORS
A. Data Set Files
Identity:
oosting_pres1.txt for the multiscale vascular plant grid data collected in 1989, with species as columns
oosting_pres2.txt contains identical data to oosting_pres1.text but has species as attributes
species_codes.txt for the species names
oosting_trees_1998.txt for the trees sampled in 1990 and 1998
oosting_env.txt for soil and altitude data collected in 1990
Size:
oosting_pres1.txt -- 1124 lines, not including the header row.
oosting_pres2.txt -- 21253 lines, not including the header row.
species_codes.txt -- 224 records, not including header row.
oosting_trees_1998.txt -- 12973 records, not including header row.
oosting_env.txt -- 289 records, not including header row.
Comments:
oosting_pres1.txt and oosting_pres2.txt:
Because grids 1 and 2 overlap (Fig. 1), they share quadrats and hence the data within these files are partially redundant. However, keeping the redundancy in the data should facilitate the utility for analysis. Data are recorded as the lowest level (smallest grain) of species occurrence. It is assumed that each species occurs at all higher levels. For example, if a certain species in the ‘Central 100’ grid was recorded in level 3 (0.5 m × 0.5 m) of a particular grid cell (see Table 1), it also is occurs in levels 4, 5, 6, 7, and 8 – but not levels 1 and 2.
oosting_trees_1998.txt:
All stems of woody species on the plot with a DBH of 2 cm or greater were censused. Species code, X and Y coordinates (nearest dm) within each of the 256-16 × 16 m grid cells, and DBH were recorded for each individual. In addition, the actual field dimensions of each grid cell were recorded (by recording corner post coordinates).
Curvature in topography, plus other errors, resulted in grid cell dimensions in the field that departed slightly from 16 × 16 m when measured parallel to the ground surface. The fluctuation in grid cell dimensions over the plot extent creates difficulty with documentation of the spatial arrangement of the tree map over the entire extent of the plot. If total plot X,Y coordinates were created using the field coordinates, there would be undesirable gaps and overlaps between cells in the resulting map. In order to provide a seamless 256 × 256 m map, raw or field coordinates (FX & F ) were adjusted to new X & Y coordinates (NX & NY) foreach grid cell so that that the final dimensions of each grid cell were 16 × 16 m. Total plot X & Y coordinates (0-256m) are reported as TX and TY.
The adjustments were made with the formulas shown below, where FX = the field X coordinate with respect to the grid cell, FY = the field Y coordinate with respect to the grid cell, NX = the new/adjusted X coordinate, NY = the new/adjusted Y coordinate. The corrected values (NX,NY) that replace the field coordinates (FX,FY) of a stem within a given grid cell are proportional to the actual field dimensions of the grid cell. For each grid cell, the corner at the origin (southwest corner) was assumed to have a right angle. Corners were numbered so that 2 refers to the location where NX=16 and NY=0, and 3 refers to the location where NX=0 and NY=16.
NX = 16 * FX * (1/X2 + (FY/(Y3 + ((FX*FY)/X2)(1/Y4 - 1/Y3)) * (1/Y4 - 1/Y3))
NY = 16 * FY * (1/Y3 + FX/(X2 + ((FX*FY)/Y3)(1/X4 - 1/X2)) * (1/X4 - 1/X2))
The data set was error checked for adjusted X,Y coordinates greater than 160 dm. Adjusted FX,FY coordinates out of range (48) were manually adjusted to fit within the standard boundaries. The following guidelines were used:
a. If 161 dm < NX or NY < 166 dm and FX or FY < 155 dm, then NX or NY = 158 dm.
b. If 161 dm < NX or NY < 166 dm and FX or FY > 155 dm, then NX or NY = 159 dm.
c. If NX or NY > 166 dm, then NX or NY = 160 dm.
Most stems fell into category b.
Post locations of each grid cell were added to the file, and adjusted and total plot coordinates were generated for these records also. There are four post location records for each grid cell. Therefore, even though only 289 posts physically exist in the field, there are 1024 (i.e., 4*256) post locations in the file. Each post has different coordinates with respect to each of the grid cells (minimum of 1, maximum of 4) it defines, so all of these records have been retained for reference.
All three sets of coordinates (Field, New, and Total plot) were retained in the file for each record. Total plot coordinates are primary; however, future sampling is likely to be done using the field coordinates in each grid cell.
Format and Storage mode: ASCII text, tab delimited. No compression schemes used.
Authentication procedures:
For the oosting_pres1.txt data, the sum of all of the species presence data is 101224. There are 21253 nonblank entries of species presence. For the oosting_pres2.txt data, the sum of all the species presence data (LEV) is 101224. In Grid 5, column 15, row 1, Carya tomentosa is present at level 3. For the species_codes.txt data, the sum of all the species id numbers is 25200. Species #102 is Ipomoea pandurata (L.) G. F. W. Meyer, and its code is IPPA. For oosting_trees_1998.txt, the sum of the 1998 diameters is 102103.55cm. Tree #9962 is Acer rubrum, with a diameter of 4.5cm in 1998. In oosting_env.txt, the sum of total exchange capacity is 3361.46. In column 15, row 5, the pH is 6.9.
B. Variable definitions
oosting_pres1.txt
Variable name |
Variable definition |
Units |
Storage type |
Range numeric values |
Missing value codes |
GRID |
Grid number (see Table 2) |
N/a |
Integer |
1–5 |
N/a |
CO |
Column or x-coordinate |
Units of 16 meters (in the case of grid numbers 1 and 2) or in units of 1 meter (in the case of grid numbers 3, 4, and 5). |
Integer |
0–15 |
N/a |
RW |
Row or y-coordinate |
Units of 16 meters (in the case of grid numbers 1 and 2) or in units of 1 meter (in the case of grid numbers 3, 4, and 5). |
Integer |
0–15 |
N/a |
Species presence (224 variables with variable headers. Codes given in species_codes.txt) |
The finest grain (i.e., lowest level) of species occurrence. |
Level ID (See table 1). |
Integer |
1–8 |
No missing data, but a blank indicates a true absence of the species |
oosting_pres2.txt
Variable name |
Variable definition |
Units |
Storage type |
Range numeric values |
Missing value codes |
GRID |
Grid number (see table 2) |
N/a |
Integer |
1–5 |
N/a |
CO |
Column or x-coordinate |
Units of 16 meters (in the case of grid numbers 1 and 2) or in units of 1 meter (in the case of grid numbers 3, 4, and 5). |
Integer |
0–15 |
N/a |
RW |
Row or y-coordinate |
Units of 16 meters (in the case of grid numbers 1 and 2) or in units of 1 meter (in the case of grid numbers 3, 4, and 5). |
Integer |
0–15 |
N/a |
CODE |
Unique Identifier for species; codes given in species_codes.txt |
N/a |
string |
n/a |
N/a |
LEV |
The finest grain (i.e., lowest level) of species occurrence. |
Level ID (See Table 1). |
Integer |
1–8 |
N/a |
species_codes.txt
Variable name |
Variable definition |
Units |
Storage type |
Range numeric values |
Missing value codes |
IDNUM |
Numeric ID Number for each species |
Number |
Integer |
1–224 |
N/a |
CODE |
Four-letter code uniquely identifying species; typically the first two letters of genus and species. |
text |
String |
N/a |
N/a |
SpeciesRAB |
Species name and authority according to Radford et al. (1968). |
text |
String |
N/a |
N/a |
SpeciesK |
Species name according to Kartesz (1999). An asterisk following the name implies some ambiguity of taxon identity or equivalence. |
text |
String |
N/a |
N/a |
oosting_trees_1998.txt
Variable name |
Variable definition |
Units |
Storage type |
Range numeric values |
Missing value codes |
ID |
Unique identifier number for each tree |
number |
integer |
1–9972 |
N/A |
CODE |
Four-letter code uniquely identifying species; codes given in species_codes.txt. Exception: “POST” means the corner post of the grid cell, not a tree |
text |
string |
N/A |
N/A |
FX |
Field-measured x-coordinate, measured as distance from W edge of the grid cell |
dm |
integer |
0–177 |
N/A |
FY |
Field-measured y-coordinate, measured as distance from S edge of the grid cell |
dm |
integer |
0–165 |
N/A |
D90 |
Diameter at Breast height 1990 |
cm |
floating point |
2–95.2 |
“.” |
C90 |
Condition Code 1990 – always 1 if observed, missing if not a tree was not observed that year. |
numeric code |
boolean |
1 |
“.” |
D98 |
Diameter at Breast height 1998 |
cm |
floating point |
0.9–91.7 |
“.” |
C98 |
Condition Code 1998 – standard condition codes for Duke Forest: 1: alive and reasonably intact 2: dead (confirmed as such by the presence of a dead stem) 3: missing (presumed dead, but no stem could be found for verification) 4: die back to below breast height. A marginal note was usually made indicating the possible cause of damage. 5: cut (sawed down by humans) 6: Damage from Hurricane Fran – see special hurricane codes, F1 through F4. Missing value: not a tree or not observed. |
number |
integer |
1–6 |
“.” |
F1 |
Uprooting status (1 = OK, 2 = partial uproot, 3 = complete uproot) |
numeric code |
integer |
1–3 |
“.” |
F2 |
Breakage (1 = less than 10% canopy loss, 2 = 10-35% canopy loss, 3 = 35-90% canopy loss, 4 = more than 90% canopy loss) |
numeric code |
integer |
1–4 |
“.” |
F3 |
Leaning (1 = OK, 2 = leaning free, with lean over 10%, 3 = supported by another tree, 4 = down (on ground) |
numeric code |
integer |
1–4 |
“.” |
F4 |
Leaned on (1 = free, 2 = upright, supporting a tree ( DBH >30% size of host), 3 = bent or leaning (crown displaced at least 10%), 4 = pinned |
numeric code |
integer |
1–4 |
“.” |
AZM |
Azimuth of trunk if it was apparently felled by Fran |
degrees |
integer |
37–340 |
“.” |
TX |
Total plot X coordinate, generated by using the adjusted X coordinate in each grid cell and adding the appropriate number of dm depending on the grid cell location |
dm |
integer |
0–2560 |
N/A |
TY |
Total plot Y coordinate, generated by using the adjusted Y coordinate in each grid cell and adding the appropriate number of dm depending on the grid cell location |
dm |
integer |
0–2560 |
N/A |
NX |
Adjusted within grid cell x-coordinate. See comments above. |
dm |
integer |
0–160 |
N/A |
NY |
Adjusted within grid cell y-coordinate. See comments above. |
dm |
integer |
0–160 |
N/A |
CL |
Indicates a clone relationship with this ID |
number |
integer |
0–9387 |
“.” |
CHECK |
Indicates by “x” a few values that are questionable and should probably be removed from analysis until validated in the field; the concerns cannot be resolved from the data sheets. |
text |
string |
N/A |
N/A |
NOTES |
Notes concerning the data, such as the nature of the possible error indicated by “CHECK”. A blank indicates no concerns. |
text |
string |
N/A |
N/A |
oosting_env.txt
Variable name |
Variable definition |
Units |
Storage type |
Range numeric values |
Missing value codes |
CO |
Column number; as in oosting_pres1.txt |
Units of 16 m |
integer |
0–16 |
N/A |
RW |
Row number; as in oosting_pres1.txt |
Units of 16 m |
integer |
0–16 |
N/A |
TEC |
Total Exchange Capacity |
MEQ/100g |
floating point |
3.77–32.65 |
N/A |
PH |
pH |
pH units |
floating point |
4.4–7.2 |
N/A |
ORG |
Organic Matter (humus) |
% |
floating point |
3–51.6 |
N/A |
S |
Soluble Sulfur |
ppm |
floating point |
14–48 |
N/A |
P |
Easily extractable Phosphorus |
ppm |
floating point |
8–36 |
N/A |
CA |
Calcium |
ppm |
floating point |
149–5706 |
N/A |
MG |
Magnesium |
ppm |
floating point |
61–461 |
N/A |
K |
Potassium |
ppm |
floating point |
30–269 |
N/A |
NA |
Sodium |
ppm |
floating point |
6–28 |
N/A |
BCA |
Saturation of calcium |
% |
floating point |
18.6–87.38 |
N/A |
BMG |
Saturation of magnesium |
% |
floating point |
6.89–27.99 |
N/A |
BK |
Saturation of potassium |
% |
floating point |
0.86–5.64 |
N/A |
BNA |
Saturation of sodium |
% |
floating point |
0.14–1.23 |
N/A |
BO |
Saturation of other cations |
% |
floating point |
4.2–8.6 |
N/A |
EXH |
Exchangable H+ |
% |
floating point |
0–57 |
N/A |
B |
Boron |
ppm |
floating point |
0.2–1.53 |
N/A |
FE |
Iron |
ppm |
floating point |
43–348 |
N/A |
MN |
Manganese |
ppm |
floating point |
71–569 |
N/A |
CU |
Copper |
ppm |
floating point |
0.61–3.77 |
N/A |
ZN |
Zinc |
ppm |
floating point |
1.1–28.6 |
N/A |
AL |
Aluminum |
ppm |
floating point |
212–1234 |
N/A |
DNS |
Density in g/cm3 |
floating point |
0.3–1 |
N/A |
|
CA/MG |
Ratio of Calcium to Magnesium |
ratio |
floating point |
1.8–21.13 |
N/A |
ALT1 |
Elevation relative to coordinate (16, 0) |
meters |
floating point |
-4.99–5.08 |
N/A |
ALT2 |
Elevation relative to minimum elevation |
meters |
floating point |
0–10.07 |
N/A |
CLASS V. SUPPLEMENTAL DESCRIPTORS
A. Data acquisition
Data forms: Data forms.
Location of completed data forms: The completed understory field data forms are stored at Oklahoma State University Department of Botany (M.W. Palmer’s Office). The completed tree and soil data forms are stored at the Department of Biology, University of North Carolina, Chapel Hill, NC 27514.
B. Quality assurance/quality control procedures. Field sheets were proofed for concerns after every day in the field.
C. Related material: n/a
D. Computer programs and data processing algorithms: n/a
E. Archiving: n/a
F. Publications and results:
These data have been used in the following publications:
Jonsson, B. G., and J. Moen. 1998. Patterns in species associations in plant communities: the importance of scale. Journal of Vegetation Science 9:327–332.
Palmer, M. W. 1995. How should one count species? Natural Areas Journal 15:124–135.
Palmer, M. W., P. Earls, B. W. Hoagland, P. S. White, and T. M. Wohlgemuth. 2002. Quantitative tools for perfecting species lists. Environmetrics 13:121–137.
Palmer, M. W., and E. van der Maarel. 1995. Variance in species richness, species association, and niche limitation. Oikos 73:203–213.
Palmer, M. W., and P. S. White. 1994. Scale dependence and the species-area relationship. American Naturalist 144:717–740.
Reed, R. A., M. W. Palmer, R. K. Peet, and P. S. White. 1993. Scale dependence of vegetation-environment correlations: a case study of a North Carolina piedmont woodland. Journal of Vegetation Science 4:329–340.
Richard, M. 1998. Fern species diversity in relation to spatial scale and structure. M.S. Thesis. McGill University, Montreal, Quebec, Canada.
Wagner, H. H. 2003. Spatial covariance in plant communities: Integrating ordination, geostatistics, and variance testing. Ecology 84:1045–1057.
Xi, W. 2005. Forest response to natural disturbance: changes in structure and diversity on a North Carolina Piedmont forest in response to catastrophic wind events. Ph.D. Dissertation, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina, USA.
G. History of data set usage: see F. above for references that utilize the data.
H. Data set update history: The vascular plant grid data were last updated in 1989. Soil data were updated in 1990. Tree data were updated in 2007. The structure of each data set was changed to its current form in 2007.
Review history: n/a
Questions and comments from secondary users: n/a
ACKNOWLEDGMENTS
M. Cassey, N. Christensen, K. Doyle, K. Jacobs, S. D. McAlister, R. McDonald, L. Meissner, I. L. Palmer, S. S. Palmer, C. Rader, M. Robinson, J. Shepherd, R. White, and K. Wolf helped in various phases of the project. We thank the following for funding: North Carolina Botanical Garden; Oklahoma State University College of Arts and Sciences; NSF grants BSR-8905926 and DEB97-07551 to RKP. MWP thanks Per Milberg of Linköpings Universitet for hosting during sabbatical leave.
LITERATURE CITED
Bowman, J., G. Forebes, and T. Dilworth. 2000. The spatial scale of variability in small-mammal populations. Ecography 23:328–334.
Bruun, H. H., J. Moen, and A. Angerbjörn. 2003. Environmental correlates of meso-scale plant species richness in the province of Harjedalen, Sweden. Biological Conservation 12:2025–2041.
Carroll, S. S., and D. L. Pearson. 1998. The effects of scale and sample size on the accuracy of spatial predictions of tiger beetle (Cicindelidae) species richness. Ecography 21:401–414.
Dennis, R. L. H. 2001. Progressive bias in species status is symptomatic of fine-grained mapping units subject to repeated sampling. Biodiversity and Conservation 10:483–494.
Heikkinen, R. K. 1996. Predicting patterns of vascular plant species richness with composite variables: a meso-scale study in Finnish Lapland. Vegetatio 126:151–165.
Huenneke, L. F., D. Clason, and E. Muldavin. 2001. Spatial heterogeneity in Chihuahuan desert vegetation: implications for sampling methods in semi-arid ecosystems. Journal of Arid Environments 47:257–270.
Jonsson, B. G., and J. Moen. 1998. Patterns in species associations in plant communities: the importance of scale. Journal of Vegetation Science 9:327–332.
Kartesz, J. T. 1999. A synonymized checklist and atlas with biological attributes for the vascular flora of the United States, Canada, and Greenland. North Carolina Botanical Garden, Chapel Hill, North Carolina, USA.
Kohler, F., F. Gillet, M. A. Progin, J. M. Gobat, and A. Buttler. 2004. Seasonal dynamics of plant species at fine scale in wooded pastures. Community Ecology 5:7–17.
Mehlich, A. 1984. Mehlich 3 soil test extraction modification of Mehlich 2 extractant. Communications in Soil Science and Plant Analysis 15:1409–1416.
NatureServe. 2007. Quercus rubra - Quercus alba - Carya glabra / Geranium maculatum Forest. Ecological association comprehensive report. NatureServe Explorer. http://www.natureserve.org/explorer/servlet/NatureServe?searchCommunityUid=ELEMENT_GLOBAL.2.683335. Observed July 7, 2007.
Newell, C. L. and R. K. Peet. 1998. Vegetation of Linville Gorge Wilderness, North Carolina. Castanea 63:275–322.
Oosting, H. J. 1956. The study of plant communities. Second edition. W.H. Freeman, San Francisco, California, USA.
Palmer, M. W. 1990. Vascular flora of the Duke Forest, North Carolina. Castanea 55:229–244.
Palmer, M. W. 1995. How should one count species? Natural Areas Journal 15:124–135.
Palmer, M. W. 2005. Distance decay in an old-growth neotropical forest. Journal of Vegetation Science 16:161–166.
Palmer, M. W., and P. M. Dixon. 1990. Small-scale environmental heterogeneity and the analysis of species distributions along gradients. Journal of Vegetation Science 1:57–65.
Palmer, M. W., P. Earls, B. W. Hoagland, P. S. White, and T. M. Wohlgemuth. 2002. Quantitative tools for perfecting species lists. Environmetrics 13:121–137.
Palmer, M. W., and G. Rusch. 2001. How fast is the carousel? Direct indices of species mobility in an Oklahoma grassland. Journal of Vegetation Science 12:305–318.
Palmer, M. W., and E. van der Maarel. 1995. Variance in species richness, species association, and niche limitation. Oikos 73:203–213.
Palmer, M. W., and P. S. White. 1994. Scale dependence and the species-area relationship. American Naturalist 144:717–740.
Peet, R. K., and N. L. Christensen. 1980. Hardwood forest vegetation of the North Carolina Piedmont. Veroffentlichungen des Geo-botanischen Institutes der Eigenossischen Technischen Hoch- schule, Stiftung Rübel 69:14–39.
Radford, A. E., H. E. Ahles, and C. R. Bell. 1968. Manual of the Vascular Flora of the Carolinas. University of North Carolina Press, Chapel Hill, North Carolina, USA.
Reed, R. A., M. W. Palmer, R. K. Peet, and P. S. White. 1993. Scale dependence of vegetation-environment correlations: a case study of a North Carolina piedmont woodland. Journal of Vegetation Science 4:329–340.
Richard, M. 1998. Fern species diversity in relation to spatial scale and structure. M.S. Thesis. McGill University, Montreal, Canada, USA.
Schafale, M. P., and A. S. Weakley. 1990. Classification of the Natural Communities of North Carolina (Third Approximation). North Carolina Natural Heritage Program, Raleigh, North Carolina, USA.
Wagner, H. H. 2003. Spatial covariance in plant communities: Integrating ordination, geostatistics, and variance testing. Ecology 84:1045–1057.
Xi, W. 2005. Forest response to natural disturbance: changes in structure and diversity on a North Carolina Piedmont forest in response to catastrophic wind events. Ph.D. Dissertation, University of North Carolina at Chapel Hill, Cahpel Hill, North Carolina, USA.